Les tissus végétaux

Préparation à l'agrégation interne SVT 2007-2008
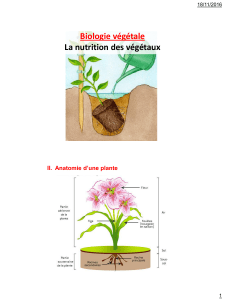
Les tissus végétaux
aspects théoriques et pratiques
Les objectifs de cette séance sont les suivants :
Manipulations :
–Réaliser une coupe et la colorer au carmino-vert de Mirande (conservation quelques années)
–Analyser des morceaux de bois (feuillus et résineux)
–détecter les réserves en amidon et protéines des tissus des graines (albumen)
–détecter l'activité amylasique de l'albumen de graines
[PRAT « L'expérimentation en physiologie végétale » Hermann éditeurs]
Analyse :
–analyser des coupes commerciales de feuilles, pétioles, tiges, racines de Mono et Dicotylédones,
dans le but de reconnaître
–(1) chaque tissu,
–(2) le type d'organe,
–(3) le type de plante,
–(4) d'éventuelles adaptations [niveau avancé]
–savoir faire et utiliser un schéma conventionnel de coupes de végétaux
Les apports théoriques serviront à comprendre les objets étudiés. Il ne s'agit pas d'un cours complet
d'anatomie de BV!
Table des matières
1.Présentation des tissus végétaux........................................................................................................3
2. Feuilles et tiges.................................................................................................................................7
3. Strucutre du bois.............................................................................................................................12
4. Exploitation du bois
5. Racine.............................................................................................................................................25

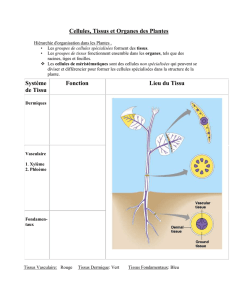
1.Présentation des tissus végétaux
Les tissus se forment à partir des méristèmes, massifs organisés de jeunes cellules indifférenciées
qui sont le siège de divisions orientées actives. Ces méristèmes peuvent être fonctionnels peu de
temps (plantes annuelles), ou pendant de nombreuses années.
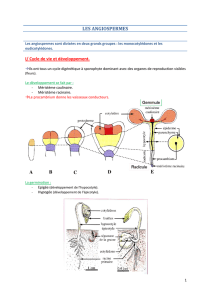
On distingue deux types de méristèmes:
- les méristèmes primaires, d'origine embryonnaire, situés à l'apex des tiges (méristèmes
caulinaires) et des racines (méristèmes racinaires), et à la base des feuilles. Ils forment les tissus
primaires qui constituent la structure primaire.
- les méristèmes secondaires, phellogène et cambium, apparaissent après les méristèmes primaires.
Ils assurent la croissance en épaisseur et donnent les tissus secondaires qui constituent la structure
secondaire.
1. Tissus de protection
L'épiderme est une assise continue de cellules qui recouvre les organes aériens et les protège contre
la dessication et les agressions extérieures tout en permettant de réguler les échanges gazeux avec
l'atmosphère. C'est un tissu vivant constitué d'une assise unique de cellules de revêtement jointives,
de cellules stomatiques et parfois de poils. L contient des cellules de revêtement, des stomates et
parfois des poils.
2. Parenchymes
Les parenchymes sont des tissus peu différenciés qui sont le siège des fonctions élaboratrices de la
plante (photosynthèse et stockage des réserves). Les cellules parenchymateuses sont en général
isodiamétriques ou allongées, plus ou moins arrondies dans les angles. Les espaces qu'elles
délimitent alors entre elles sont appelés méats ou lacunes selon leur taille.
- parenchyme chlorophylliens ou assimilateurs, palissadique ou lacuneux (Dicotylédones)
- parenchymes de réserve
- rôle dans la mise en réserve de matériaux élaborés par les cellules chlorophylliennes,
- présents dans de nombreux organes tels que les racines, les tiges souterraines (ou
rhizomes), les tubercules, les tiges aériennes, les fruits et les graines.
- parenchyme aquifère, qui contient de l'eau chez les plantes succulentes
- parenchyme aérifère qui contient de l'air chez les plantes aquatiques.
Dans les organes âgés, souvent le parenchyme devient sclérifié par lignification des parois de
cellules parenchymateuses dans des organes dont la croissance est achevée.
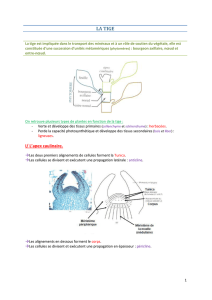
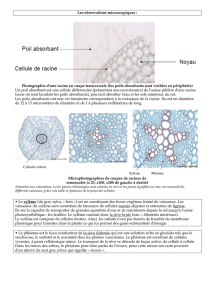
3. Tissus conducteurs
Chez les Angiospermes, les gymnospermes et les fougères, la circulation des sèves est assurée par
un appareil conducteur composé de deux types de tissus : le xylème et le phloème.
Xylème
Le xylème des Angiospermes contient trois types d'éléments : des fibres de type trachéide qui
assurent le soutien, des cellules de parenchyme et des vaisseaux qui assurent la conduction. Selon le
dessin de la lignification on distingue les vaisseaux : annelés, spiralés, rayés, réticulés, ponctués.
Les cellules parenchymateuses dites "de contact" qui bordent les vaisseaux assurent la sécrétion des
ions dans le xylème ou des sucre au début du printemps (sève brute d'érable = sirop d'érable).
Phloème
Le phloème, ou liber, conduit la sève élaborée, solution de substances organiques riches en glucides,
des feuilles vers les autres organes.

LE
PHLOÈME
éléments conducteurs
(tissu vivant)
- cellules allongées disposées en file
communiquant entre elles par des pores qui
interrompent la paroi cellulosique,
- pas de noyau ni de vacuole.
cellules compagnes - bordent les tubes criblés.
cellules
parenchymateuses - de réserve, sécrétrices.
fibres - cellules à paroi épaisse, lignifiées ou
cellulosiques.
4.Tissus de soutien
Ils peuvent être pectocellulosiques (=roses) comme le collenchyme, ou ligneux (=verts) comme le
sclérenchyme à parois épaisses, le parenchyme sclérifié à paroi fines et le xylème II au centre de la
tige (bois de coeur).
Les tissus de soutien assurent souplesse et rigidité aux organes de la plante. Le collenchyme se
forme dans les organes jeunes tandis que le sclérenchyme se rencontre dans les organes dont
l'allongement est achevé.
LE COLLENCHYME
Le collenchyme se forme dans les organes jeunes en croissance, aériens essentiellement. C'est un
tissu vivant dont les parois sont épaissies par un dépôt de cellulose, ce qui confère à la plante une
grande résistance à la flexion et à la traction, une élasticité et une certaine souplesse. Il est
généralement situé en anneaux ou en îlots sous l'épiderme des tiges et des pétioles, ou encore accolé
à des vaisseaux conducteurs dans les pétioles ou les limbes des feuilles.
LE SCLÉRENCHYME
Le sclérenchyme est le tissu de soutien des organes dont l'allongement est achevé. C'est un tissu
constitué de cellules mortes dont les parois sont épaissies par un dépôt de lignine qui confère dureté
et rigidité à la plante.

Structure moléculaire de la cellulose
Par suite de la conformation de la molécule de cellobiose, les liaisons osidiques successives (β 1-4)
ainsi que les fonctions CH2OH sont situées alternativement au dessus et au dessous du plan de la
molécule. Cette conformation permet de solidifier cette chaîne par des liaisons hydrogène ( liaisons
H) répétitives. Grâce à cela, la molécule présente une forme linéaire et ne s'enroule pas sur elle-
même. En fait, les molécules de cellulose ne sont pas libres mais sont associées entre elles par des
liaisons H inter-caténaires pour constituer des microfibrilles extrêmement résistantes. La cellulose,
associée aux pectines, représente le constituant majeur des parois primaires en assurant à la fois le
solidité et une certaine élasticité.
En présence de protons, les liaisons H sont détruites, ce qui permet l'élongation de la paroi des
jeunes cellules.
Dans les parois secondaires, la cellulose forme des macrofibrilles. Les macrofibrilles de cellulose en
grande quantité, comme dans le collecnhyme, assurent un rôle de soutien.

La lignine
La lignine (ou plutôt les lignines) est un polymère complexe, non linéaire, constitué de monomères
constitués d'un squelette phenyl-propane (cycle de 6 carbones . chaîne à 3 carbones)
Les trois monomères principaux de la lignine.
L'acide conyférylique est très abondant (80%) chez les Conifères (Gymnospermes) et l'acide
sinapylique (50%) chez les Angiospermes.
La polymérisation n'est pas linéaire et les motifs de base associés entre eux constituent un réseau
qui s'interpénètre dans le réseau de cellulose. Contrairement à ce que l'on pourrait penser, ce n'est
pas la paroi "secondaire" des éléments du bois qui est la plus lignifiée mais plutôt la partie
mitoyenne entre les cellules (lamelle moyenne), ce qui renforce la cohésion et donc la résistance
mécanique du bois.
Ceci a plusieurs conséquences :
–Dans la plante, l'hydrophobie de la lignine est un élément majeur de la bonne circulation de la
sève
–Le bois coupé et séché (matériau) a une densité inférieure à 1. Placé dans l'eau, il continue de
flotter car il ne s'imbibe pas d'eau grâce à la lignine.
Les produits de dégradation de la lignine sont un des éléments majeurs de l'humus.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%