les principales espèces de charadriiformes sur la vistule moyenne

Dariusz Bukaciński Monika Bukacińska
LES PRINCIPALES ESPÈCES
DE CHARADRIIFORMES
SUR LA VISTULE MOYENNE
LA BIOLOGIE, L’ÉCOLOGIE, LA CONSERVATION
ET L’OCCURRENCE
ISBN 978-83-941734-2-5
STERNE NAINEGOÉLAND CENDRÉ
STERNE PIERREGARIN GRAND GRAVELOT
MOUETTE RIEUSE
PETIT GRAVELOT
LES MONOGRAPHIES
De la part des auteurs
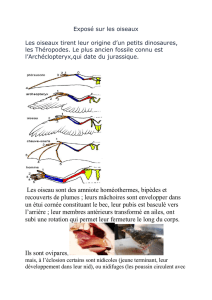
Entre vos mains vous tenez des monographies de six espèces d’oiseaux pour lesquels la vallée
de la Vistule moyenne est un site de nidification très important dans le pays. Nous avons essayé
de vous transmettre des connaissances leur concernant d’une manière accessible, tout en
n’ignorant pas les aspects importants de leur biologie et écologie, en particulier les questions
liées au comportement de reproduction. L’écologie comportementale : la recherche des causes,
des conditions et des conséquences des tactiques et des stratégies de reproduction utilisées
par les oiseaux – est notre spécialité professionnelle. Dans la présente publication, nous avons
également inclu des informations inédites de nos recherches dans les colonies de la Vistule
des charadriiformes. Pour la plupart, elles se rapportent à l’adaptation comportementale des
conditions extrêmement dynamiques et instables d’un grand fleuve de pleine, tressé, qu’est la
Vistule dans son cours moyen.
Les monographies en français sont une version abrégée des monographies en polonais.
Le projet « Protection des habitats d’espèces clés d’oiseaux dans la Vallée de la Vistule Moyenne dans des
conditions de pression intense de l’agglomération de Varsovie » (WislaWarszawska.pl) a été cofinancé dans le
cadre de l’instrument financier LIFE+ de la Communauté européenne et du Fonds National pour la Protection
de l’Environnement et la Gestion de l’Eau.

Dariusz Bukaciński Monika Bukacińska
LES PRINCIPALES ESPÈCES
DE CHARADRIIFORMES
SUR LA VISTULE MOYENNE
LA BIOLOGIE, L’ÉCOLOGIE, LA CONSERVATION
ET L’OCCURRENCE
LES MONOGRAPHIES
Varsovie 2015

Titre original:
Kluczowe gatunki ptaków siewkowych na środkowej Wiśle: biologia, ekologia, ochrona i występowanie.
Monografie
La revue scientifique
dr hab. Patryk Rowiński
Texte, tableaux, graphiques
Dariusz Bukaciński, Monika Bukacińska
Institut de l’écologie et de bioéthique, Faculté de philosophie chrétienne
Université Cardinal Stefan Wyszyński de Varsovie
Rédacteur en chef
Ewa Kominek, Małgorzata Zygmont
Rédaction, correction
Adam Chęć (Partie 1–4), Agnieszka Łobik-Przejsz (Partie 5–6), Małgorzata Zygmont
Traduction
besttext.pl
Les photographies
sur la couverture: Dariusz Bukaciński – le paysage, Ewa Kominek – Goéland cendré, Mouette rieuse,
Marcin Łukawski – Petit Gravelot, Maciej Szymański – Sterne pierregarin, Grand Gravelot, Artur Tabor – Sterne naine
dans le texte: Jakub Badełek – p. 95; Arkadiusz Buczyński - p. 8 en haut à gauche, 9 en bas, 10 en haut, 11 en
bas à gauche, 25 à droite, 28 à droite, 29, 30 en bas, 31, 32 à droite, 50, 51, 52 au milieu, 55, 64 en bas, 65, 66,
67 en haut et en bas, 82 en haut, 84 à gauche, 85, 86, 90, 105, 107, 108, 109 en haut et en bas à droite;
Dariusz Bukaciński – p. 8 en haut à droite et en bas, 9 en haut à gauche, 10 en bas, 11 en haut et au milieu
à droite, 15, 28 à gauche, 52 en haut et en bas, 64 en haut, 67 au milieu, 82 au milieu et en bas, 84 à droite,
109 en bas à gauche; Leszek Iwanowski de la société Hydrolech – p. 73, 80, 93; Grzegorz Leśniewski – p. 11 au
milieu à gauche, 87; Marcin Łukawski – p. 23, 37, 48, 49, 56, 62; Marek Sawicki – p. 9 en haut à droite, 11 en
bas à droite, 22, 25 à gauche, 30 en haut, 32 à gauche, 35, 102 en bas, 103 à droite, 111, 113, 114;
Maciej Szymański – p. 7, 103 à gauche; Artur Tabor – p. 6, 102 en haut
Cartes
Alina Gerlée
Conception graphique, mise en page, impression
Lotos Poligrafia Sp. z o.o., www.lotos-poligrafia.pl
© Copyright: Ville capitale de Varsovie. All rights reserved.
Varsovie 2015
ISBN 978-83-941734-2-5
La publication a été émise par l’Association de Varsovie pour la Protection des Oiseaux dans le cadre du projet
« Protection des habitats d’espèces clés d’oiseaux dans la Vallée de la Vistule Moyenne dans des conditions
de pression intense de l’agglomération de Varsovie » (WislaWarszawska.pl), cofinancé de l’Instrument finan-
cier LIFE+ de la Communauté européenne et du Fonds national pour la Protection de l’Environnement et de la
Gestion de l’Eau.
Les monographies en français sont une version abrégée des monographies en polonais.
Stowarzyszenie Stołeczne Towarzystwo Ochrony Ptaków
(STOP; L’Association de Varsovie pour la Protection des Oiseaux)
Czeska 15a/5, 03-902 Varsovie, www.stop.eko.org.pl
La méthode recommandée pour citer l’ouvrage
Bukaciński D., Bukacińska M. 2015. Les principales espèces de Charadriiformes sur la Vistule moyenne : la bio-
logie, l’écologie, la conservation et l’occurrence. Les monographies. L’Association de Varsovie pour la Protection
des Oiseaux (STOP), Varsovie, 138 p.

3
TABLE DES MATIÈRES
PARTIE 1. GOÉLAND CENDRÉ Larus canus. MONOGRAPHIE
Goéland cendré Larus canus ........................................................ 6
Caractéristiques de l’espèce, mode de vie ............................................ 6
Milieu de vie ....................................................................... 7
Biologie de la reproduction.......................................................... 8
Modèles sociaux et comportements reproducteurs ................................... 12
Migration et hivernage.............................................................. 14
Nourriture ......................................................................... 14
État de la population, menaces et protection ......................................... 16
Repartition et abondance en Pologne ................................................ 16
Menaces et protection en Pologne................................................... 17
PARTIE 2. MOUETTE RIEUSE Chroicocephalus ridibundus. MONOGRAPHIE
Mouette rieuse Chroicocephalus ridibundus .......................................... 22
Caractéristiques de l’espèce, mode de vie ............................................ 22
Milieu de vie ....................................................................... 24
Biologie de la reproduction.......................................................... 25
Modèles sociaux et comportements reproducteurs ................................... 34
Migration et hivernage.............................................................. 40
Nourriture ......................................................................... 41
État de la population, menaces et protection ......................................... 42
Repartition et abondance en Pologne ................................................ 43
Menaces et protection en Pologne................................................... 44
PARTIE 3. STERNE NAINE Sternula albifrons. MONOGRAPHIE
Sterne naine Sternula albifrons ..................................................... 48
Caractéristiques de l’espèce, mode de vie ............................................ 48
Milieu de vie ....................................................................... 49
Biologie de la reproduction.......................................................... 50
Modèles sociaux et comportements reproducteurs ................................... 53
Migration et hivernage.............................................................. 56
Nourriture ......................................................................... 56
État de la population, menaces et protection ......................................... 57
Repartition et abondance en Pologne ................................................ 58
Menaces et protection en Pologne................................................... 58

4
PARTIE 4. STERNE PIERREGARIN Sterna hirundo. MONOGRAPHIE
Sterne pierregarin Sterna hirundo ................................................... 62
Caractéristiques de l’espèce, mode de vie ............................................ 62
Milieu de vie ....................................................................... 63
Biologie de la reproduction.......................................................... 65
Modèles sociaux et comportements reproducteurs ................................... 68
Migration et hivernage.............................................................. 71
Nourriture ......................................................................... 72
État de la population, menaces et protection ......................................... 74
Repartition et abondance en Pologne ................................................ 75
Menaces et protection en Pologne................................................... 76
PARTIE 5. PETIT GRAVELOT Charadrius dubius. MONOGRAPHIE
Petit Gravelot Charadrius dubius .................................................... 80
Caractéristiques de l’espèce, mode de vie ............................................ 81
Milieu de vie ....................................................................... 82
Biologie de la reproduction.......................................................... 83
Modèles sociaux et comportements reproducteurs ................................... 88
Migration et hivernage.............................................................. 94
Nourriture ......................................................................... 95
État de la population, menaces et protection ......................................... 96
Repartition et abondance en Pologne ................................................ 96
Menaces et protection en Pologne................................................... 98
PARTIE 6. GRAND GRAVELOT Charadrius hiaticula. MONOGRAPHIE
Grand Gravelot Charadrius hiaticula ................................................. 102
Caractéristiques de l’espèce, mode de vie ............................................ 103
Milieu de vie ....................................................................... 105
Biologie de la reproduction.......................................................... 106
Modèles sociaux et comportements reproducteurs ................................... 111
Migration et hivernage.............................................................. 116
Nourriture ......................................................................... 117
État de la population, menaces et protection ......................................... 118
Repartition et abondance en Pologne ................................................ 119
Menaces et protection en Pologne................................................... 120
CONCLUSION ......................................................................... 122
LITTÉRATURE DE RÉFÉRENCE ........................................................... 123
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
1
/
140
100%