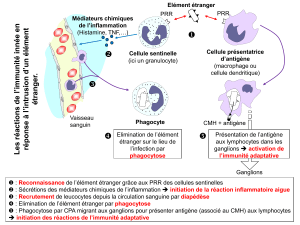
immunite a mediation cellulaire

Immunité à médiation cellulaire (3
ème
année Biochimie) Dr Belambri S. A
Immunité à médiation Cellulaire
Introduction :
Une fois leur développement terminé dans le thymus, les cellules T pénètrent dans la
circulation sanguine, transportées dans le flux sanguin où elles seront en perpétuelles recirculation
entre le sang et les tissus lymphoïdes périphériques jusqu’à ce qu’elles rencontrent leurs antigène
spécifique. Les cellules T matures circulantes qui n’ont pas encore rencontré leur antigène sont dites
cellule T naïves. Leur rencontre avec l’antigène induit son activation et donc sa prolifération et
différentiation en cellule T effectrice armée. Les cellules sur lesquelles les cellules T effectrices
armée agissent sont appelées cellules cibles.
Les cellules T naïves sont activées lorsque les CPA, principalement les cellules dendritiques
leur présentent l’antigène sous forme de complexe CMHII-peptide à leur surface. Les macrophages
et les cellules B peuvent aussi se comporter comme des CPA. Les cellules T effectrices sont
regroupées en trois classes fonctionnelles qui peuvent détecter les peptides antigéniques provenant
de différents types de pathogènes (figure 1) :
- Les peptides provenant de pathogènes se multipliant dans le cytoplasme sont associés aux
molécules de CMH de classe I et sont exprimés à la surface aux cellules T CD8+ qui se
différentient en cellules T cytotoxiques capable de tuer la cellule cible infectée.
- Les peptides qui proviennent soit des pathogènes qui se multiplient dans les vésicules
intracellulaires soit des bactéries et toxines extracellulaires capturées par les cellules sont
transportés à la surface et exprimés en association avec les CMH-II et présentés aux cellules T
CD4+. Ces cellules T CD4+ peuvent se différentier en deux types de cellules T effectrices : les
cellules Th1 et Th2.
- Les pathogène qui s’accumulent en grand nombre dans les macrophages et dans les vésicules des
cellules dendritiques ont tendance à stimuler la différentiation des cellules Th1. Ces cellules
activent les propriétés microbicides des macrophages et stimulent les cellules B à produire des IgG
permettant d’opsoniser les pathogènes extracellulaires favorisant leur capture par les phagocytes.
- Les antigènes extracellulaires ont tendance à stimuler la production des cellules Th2 qui sont à
l’origine de la réponse humorale en stimulant la production des IgM par les cellules B naïves
spécifiques de l’antigène et par la suite la production des autres isotypes d’anticorps (IgA, IgE) ainsi
que l’IgG neutralisant et/ou faiblement opsonisant.
Ainsi l’activation des cellules T naïves par l’antigène ainsi que la prolifération et la
différentiation qui en découlent constituent la réponse immunitaire primaire à médiation
cellulaire qui fournit des cellules T effectrices et mémoires pour une protection lors de la rencontre
ultérieur avec le même antigène.

Immunité à médiation cellulaire (3
ème
année Biochimie) Dr Belambri S. A
Figure 1. Rôle des cellules T effectrices dans les réponses de types cellulaire et humorale contre les
pathogènes les plus typiques.
1. Production des cellules T effectrices armées
Pour être activée, une cellule T naïve doit reconnaître un peptide étranger fixé à une
molécule du CMH du soi. Mais cette reconnaissance n’est pas suffisante à elle seule, car
l’activation des cellules T nécessitent en même temps un signal de co-stimulation fournit par une
CPA. Seule les cellules dendritiques, les macrophages ou encore les cellules B peuvent exprimer à
la fois les molécules du CMH et les molécules de co-stimulation qui permettent l’expansion clonale
des cellules T naïves et leur différentiation en cellules effectrices armées.
Il est important de noter que les réponses immunes adaptives ne prennent pas naissance à
l’endroit où le pathogène établit le premier foyer infectieux. En fait, elles ont lieu dans les tissus
lymphoïdes périphériques à travers lesquels les cellules T naïves circulent continuellement. Le
pathogène ou leurs composants sont transportés dans les tissu lymphoïdes par la lymphe qui draine
les tissus infectés, ou plus rarement par le sang. Ainsi par exemple, les pathogènes qui infectent les
surfaces des muqueuses s’accumulent dans les tissus lymphoïdes comme les plaques de Payer et les
amygdales. Ceux qui pénètrent par le sang sont piégés dans la rate alors que ceux qui infectent les
tissus périphériques sont piégés dans les ganglions lymphatiques. Comme on l’a vu dans le chapitre
1, tous ces tissus lymphoïdes possèdent des cellules spécialisées dans la capture et la présentation
des antigènes aux cellules T dont les plus importantes sont les cellules dendritiques qui capturent

Immunité à médiation cellulaire (3
ème
année Biochimie) Dr Belambri S. A
l’antigène sur le site de l’infection puis migrent vers les ganglions lymphatiques. Les cellules
dendritiques peuvent capturer, apprêter et présenter une grande variété de pathogènes et
d‘antigènes. Elles semblent être les activatrices des cellules T naïves les plus importantes. Les
macrophages et les cellules B semblent être spécialisés dans l’apprêtement et la présentation des
respectivement des pathogènes phagocytés et antigènes solubles (toxines) (Figure 2).
Figure 2. Les cellules présentatrices d’antigène sont présentes dans différentes parties du
ganglion.

Immunité à médiation cellulaire (3
ème
année Biochimie) Dr Belambri S. A
Les cellules T naïves pénètrent dans le tissu lymphoïde en traversant les parois des veinules
connues sous le nom de veinules à endothélium épais (HEV) (Figure 3). Continuellement ces
cellules passent de la circulation sanguine aux organes lymphoïdes puis retournent dans le sang,
entrant ainsi chaque jour en contact avec plusieurs milliers de cellules présentatrices de l’antigène
dans les tissus lymphoïdes. Les cellules qui n’ont pas rencontré leur antigène spécifique atteignent
la zone médullaire du ganglion lymphatique à partir de laquelle elles sont renvoyées dans la
circulation sanguine via les vaisseaux lymphatiques. On estime que seule une cellule T naïve sur
10
4
à 10
6
a une chance de reconnaître un antigène précis et l’immunité adaptive dépend de
l’expansion (prolifération) de ces rares cellules spécifiques de l’antigène (figure 3), ces cellules
arrêtent alors de migrer et commencent les différentes étapes qui vont les conduire à la
différentiation en cellules effectrice armées. En effet, après plusieurs jours, les cellules effectries
armées qui se sont développées à partir d’une cellule T naïve quittent le ganglion lymphatique et
retournent dans le sang pour se diriger vers le site de l’infection.
Figure 3. Les cellules T naïves rencontrent leur antigène spécifique lors de leur passage dans les
organes lymphoïdes périphériques.

Immunité à médiation cellulaire (3
ème
année Biochimie) Dr Belambri S. A
2. Stimulation de l’expansion des cellules T naïves
Les cellules T naïves sont activées lorsque leur TCR et leur co-récepteur (CD4 ou CD8) se
fixe au complexe CMH-peptide à la surface des CPA. Cependant, l’expansion clonale de ces
cellules T ayant reconnu le complexe CMH-peptide nécessite un deuxième signal ou signal de co-
stimulation qui est fourni par la même cellule qui lui a présenté le complexe CMH-peptide. Les
molécules de co-stimulation les mieux caractérisées sont les molécules B7 (B7.1 et B7.2). Ces
molécules sont des homodimères de la famille des immunoglobulines exprimées uniquement à la
surface des cellules qui peuvent stimuler la prolifération des cellules T et reconnaissent le CD28 à la
surface de celle-ci (figure 4). Cependant, une fois qu’une cellule T naïve a été activée, elle exprime
un grand nombre de protéines à sa surface qui lui permettent de maintenir ou de modifier le signal
de co-stimulation qui a induit l’expansion clonale et la différentiation. Une de ces protéines est le
CD40L qui se lie au CD40 à la surface des CPA. Ainsi la liaison du CD40L au CD40 envoie des
signaux d’activation aux cellules T et en même temps stimule les CPA à exprimer plus de
molécules B7 pour stimuler encore plus la prolifération des cellules T. Un autre couple de
molécules semble contribuer à la co-stimulation des cellules T, à savoir les molécules 4-1BB qui se
fixe à son ligand 4-1BBL à la surface des cellules dendritiques. L’interaction de ces deux molécules
active aussi bien les cellules T que les cellules dendritiques. Par ailleurs, après activation des
cellules T, elles expriment à leur surface un autre ligand des molécules B7 qui est le CTLA-4 qui
présente une affinité au B7 20 fois supérieure à celle du CD28. La liaison CTLA-4-B7 au lieu de
CD28-B7 rend la cellule T activée moins réceptive aux signaux envoyés par la CPA, ce qui conduit
à l’inhibition de sa prolifération.
Ainsi, il apparait que les cellules présentatrices de l’antigène engagent un dialogue de co-
stimulation avec les cellules T qui reconnaissent l’antigène qu’elles lui présentent. Ce dialogue
semble être initié par la fixation du B7, exprimé à la surface de la CPA activée, au CD28 de la
cellule T naïve.
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%