– ue7: –

2016-2017 Paroi bactérienne
Biologie des procaryotes et des virus
– ue7: –
Paroi bactérienne
Il n'y a pas d'annexe
Semaine : n°1 (du 05/09/16 au
09/09/16)
Date : 08/09/2016
Heure : de 16h00 à
18h00 Professeur : Pr. Romond
Binôme : n°13 Correcteur : n°32
Remarques du professeur aucune remarque
PLAN DU COURS
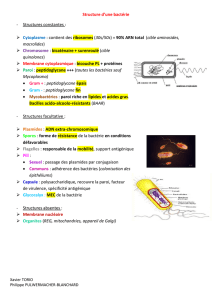
I) Rappels sur la structure et la composition du peptidoglycane
A) Points à retenir dans la constitution du peptidoglycane
B) Biosynthèse du peptidoglycane
II) Antibiotiques qui agissent sur la paroi bactérienne
A) Penicilline
B) Vancomycine
C) Antibiotiques qui vont viser une partie de la biosynthèse des bactéries
III) Mycobactéries
A) Paroi spécifique des mycobactéries
B) Composition
IV) L'ouverture de septum
A) Observation de la bactérie escherichia e coli
B) Observation de la bactérie staphylococcus aureus
C) Observation de la bactérie streptomyces venezuelae
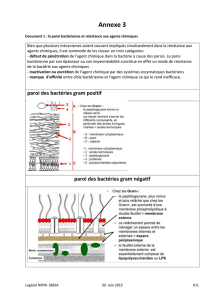
V) Eléments distinctifs entre GRAM +, GRAM – et les mycobactéries
A) Éléments spécifiques aux GRAM +
B) Eléments spécifiques aux GRAM -
C) Paroi de mycobactérie
VI) L'appareil nucléaire
A) Les chromosomes
B) Les plasmides
1/14

2016-2017 Paroi bactérienne
C) Les ribosomes
VII) La spore
A) La structure de la spore
B) Les constituants
C) Le cycle sporal
1) La sporulation
2) La germination
2/14

2016-2017 Paroi bactérienne
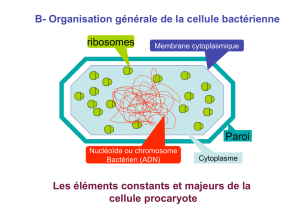
I) Rappels sur la structure et la composition du peptidoglycane
A) Points à retenir dans la constitution du peptidoglycane :
Le peptidoglycane est composé :
–D'une chaine oligosaccharridique avec une alternance de N-Acétyl Glucosamine (NAG) et de N-Acétyl
Muramique (NAM) reliés par des liaisons β 1-4.
–D'une chaine peptidique formée de 4 acides aminés alternant les formes L et D (sachant que ce sera
toujours une D-Alanine en 4e position) avec une présence possible d'acide diaminopimelique (DAP).
Cette chaine peptidique effectue toujours un branchement sur le N-Acétyl Muramique.
Dans un feuillet : les liaisons inter-peptidiques se font de sorte que le 4e acide aminé de la chaine
peptidique n°1 sera relié vers le 3e acide aminé de la chaine peptidique suivante. On a donc une D-Alanine
sur deux qui est bloquée.
La liaison entre la 1ère et la 2e chaine peut se faire par lien direct mais parfois, le pont inter-peptidique
peut être beaucoup plus varié avec 2, 3, 4 acides aminés qui ne seront pas de même nature.
C'est donc en ayant un système alterné de liaisons oligosaccharidiques que l'on va créer un feuillet
B) Biosynthèse du peptidoglycanne
C'est dans le cytosol que l'on a les précurseurs
On part de la N-Acétyl Glucosamine et avec un phosphoénolpyruvate, on va créer la N-acétyl Muramate.
Ensuite un premier L-Alanine va venir se brancher.
Puis de la glutamine vient se fixer et ensuite on aura 2 possibilités de présentées (non exclusives)
–soit on rajoute une lysine
–soit on rajoute un DAP
Les bactéries peuvent avoir une seule chaine peptidique pour l'ensemble du protéoglycane mais aussi 2 voire 3
chaines oligosaccharidiques pour un seul peptidoglycane.
Cela veut dire qu'on aura des pourcentages de chaines pour l'ensemble du peptidoglycane.
ATTENTION : Cette composition ne reste qu'un exemple !
Ensuite on rajoute 2 D-Alanines finaux et une fois que tout ceci est formé c'est ce qu'on appelle le nucléotide de
Park.
Une fois qu'on a ce nucléotide de Park , ce qui nous importe c'est d'avoir une liaison avec un lipide pour passer la
membrane. Donc on va utiliser l'undecaprénol que l'on va charger en phosphate pour qu'il puisse rendre notre
système plus hydrophobe.
Cependant il nous faut encore de la N-acétylglucosamine pour recréer une unité de peptidoglycane.
Et ensuite on pourra passer la membrane plasmique et libérer le Undecaprényl phosphate (UDP) qui va se
retrouver de l'autre coté de la membrane, pour qu'il puisse se régénérer et repartir avec un autre nucléotide de Park.
De même qu'à ce moment il y aura l'addition des différentes chaines inter- peptidiques si besoin mais dans le
système de base, on a besoin d'une seule liaison directe entre deux chaines peptidiques
On aura donc fait passer de l'autre côté de la membrane :
–le NAG,
–le NAM
–le lipide
–et 4 acides aminés.
3/14

2016-2017 Paroi bactérienne
Et à ce moment là on aura l'addition des chaines inter-peptidiques si besoin (cela va se passer avant qu'on utilise
les enzymes qui sont des transpeptidases, des carboxypeptidases et transglycosylases qui vont greffer cette
nouvelle unité sur le peptidoglycane « ouvert »)
II) Antibiotiques qui agissent sur la paroi bactérienne
•Les carboxypeptidases et transpeptidases permettent de supprimer le 5e D-Alanine qui va être perdu au
moment de la transpeptidation.
•La transpeptidase sert à faire la liaison avec le 3e acide aminé (ou son équivalent si c'est une chaine inter-
peptidique)
•La transglycosylase a pour but de lier le disaccharide (NAG et NAM) à l'ancienne chaine
A) La pénicilline
Les transpeptidases et les carboxypeptidases vont être connues sous le terme de PLP : protéines liants la
pénicilline.
Le cycle β-lactane peut se fixer sur ces deux enzymes et comme cela forme une liaison covalente et stable, il n'y
aura pas de possibilité que ces enzymes soient actives. Elle ne pourront donc pas libérer le 5e acide aminé et cela
va donc bloquer la capacité de transpeptidation.
Le principe de base de la pénicilline est d’empêcher la formation de la paroi (mais attention cela n’empêche
pas la formation de la membrane plasmique).
Il n'y aura donc pas d'allongement de la bactérie ni de création de septum. On aura un patchwork de l'ancienne
paroi avec la membrane plasmique uniquement, et lorsque cette membrane sera dans un milieu néfaste pour elle,
elle se détruira.
De ce fait, la pénicilline ralentit considérablement la prolifération bactérienne.
B) La vancomycine
La vancomycine et la teicoplanine (???) , fonctionnent de la même façon, le 5e D-Alanine qui se trouve dans la
partie péri-plasmique va être bloqué par la vancomycine de l'extérieur. Et dès lors qu'ils seront liés, il n'y aura plus
du tout d'activité possible avec d'autres unités. Donc on ne peut pas faire de transoxydation ni de transpeptidation
et on ne peut pas couper ce 5e D-Alanine.
On parle de résistance par modification de PLP car si on a une bactérie qui modifie ces séquences, il n'y aura
plus de reconnaissance des structures β-Lactanes et la bactérie continue de se diviser. La résistance vient du fait
qu'on peut avoir des mutations sur les PLP.
C'est la même chose au niveau de la vancomycine, car il peut y avoir une modification au moins d'un des types
d'enzymes pour que cela suffise à produire encore de la paroi
Chez les bactéries, il n'y a pas une séquence unique de transpeptidases, ou une séquence unique de
carboxypeptidases mais il y en a toute une série de séquences, déjà naturellement présentes mais en plus on pourra
trouver des mutations qui vont apparaître par sélection.
C) Antibiotiques qui vont viser une partie de la biosynthèse de la bactérie
•La Fosfomycine : Elle va inhiber la synthèse du nucléotide de park. A partir du moment où on n'a pas la
capacité à partir du précurseur de faire une liaison entre le NAM et les acides aminés, le reste ne
fonctionnera pas.
•La Bacitracine : Elle inhibe la phosphorylation de l'undécaprénol. Or sans lipide on ne peut pas passer
la membrane plasmique donc ça va rester au centre du cytosol.
4/14

2016-2017 Paroi bactérienne
Synthèse :
–On a d'abord dans le cytosol les précurseurs qui vont former le nucléotide de Park . Ce qui est important
c'est qu'il y a les deux DLA terminaux qui vont former le nucléotide de Park.
–Dès lors qu'il est formé ,on a besoin de l'Undecaprényl phosphate qui va régir , pour réussir à recréer une
nouvelle liaison entre le nucléotide Park et un Undecaprényl, l'ensemble s'appelant lipide 1
–Quand on aura ajouté la NAG , à ce moment on aura le lipide 2
–Cela va passer dans l'environnement direct de la membrane plasmique parce que l'on a pas de lipide
flottant dans le cytoplasme
–On a donc translocation de la membrane plasmique et donc perte de l' Undecaprényl phosphate (UDP)
–Il y a aussi une ouverture du peptidoglycane avec soit un lysozyme ou une muraminidase : deux enzymes
qui vont ouvrir la chaine oligosaccharridique
–Les transpeptidases et carboxypeptidases vont ouvrir la chaine peptidique
–Et on va avoir besoin des transpeptidases et des transglycosylases pour lier cette nouvelle unité à
l'ancienne chaine de protéoglycanes
–Ces enzymes sont cibles de la pénicilline
III) Mycobactéries
2 mycobactéries connues :
Mycobacterium Tuberculosis
–Mycobactérium Leprae
A) La paroi spécifique des mycobactéries
La coloration de Gram permet de voir les bactéries Gram + et Gram – mais ici vu la composition particulière de
la paroi, on va se retrouver en incapacité de faire diffuser les colorants à froid à l'intérieur de la cellule
On va utiliser une autre approche, une coloration à chaud : coloration de Ziehl qui est dédiée aux mycobactéries,
pour réussir à passer à travers leur paroi, on va faire bouillir le colorant sur la lame et on aura une première phase
acido-alcolique pour réussir à éliminer une partie de ces lipides.
ATTENTION : Nous sommes encore au niveau du peptidoglycane , pas encore sur l'ensemble de la paroi ,
donc nous allons voir que l'aspect protéoglycanique
Par rapport à la structure des peptidoglycanes vue précédemment, celle des mycobactéries va différer
On va avoir une structure avec une chaine qui commence par une NAG suivie par un rhamnose, et ensuite on va
trouver cette fameuse chaine composée de galactofuranoses, laquelle est branchée par des arabino furanoses.
Ce n'est pas cette structure qui est hydrophobe mais ce seront les glycolipides qui s'accrocheront grâce aux sucres.
B) Composition
La chaîne oligosaccharidique est un peu différente, l'acide N-acétyl muranique (NAM) va être remplacé par de
l'acide N glycosyl muranique (NGM). On a une chaîne de sucres.
On a une membrane plasmique qui est du côté du cytosol. Au début on aura une structure de type peptidoglycane.
Jusque là, pas de grosses différences. On met le peptidoglycane. On va voir du lipoarabino-mannane que l'on
avait pas avant. On va voir des chaînes que le peptidoglycane traverse. On va avoir aussi des structures arabino-
galactanes qui viennent s'enchâsser sur la partie peptidoglycane. Au dessus, on va retrouver des acides
mycoliques. On les a en chaînes. Et on va voir aussi des glycolipides phénoliques.
Donc la chaine peptidoglycanique est la même, mais sur la partie extérieure qui se retrouve directement avec
l'environnement de la bactérie, on aura des chaines glycaniques . C'est pourquoi on n'aura pas les mêmes
sensibilités.
5/14
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%