NB - P3C4 - Syst me moteur

NEUROBIOLOGIE!
PARTIE 3: SOMESTHÉSIE, VISION, AUDITION, SYSTÈME MOTEUR (GRAMMONT)!
!
CHAPITRE 4: LE SYSTÈME MOTEUR
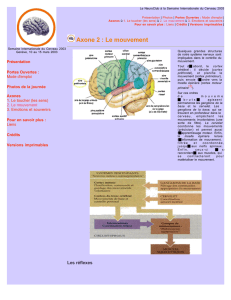
➤ L’action: logique et constitution du système nerveux moteur!
•Si le système sensoriel suivent plus ou moins une logique ascendante, on pourrait croire
que l’on suive une logique descendante lorsqu’on traite des systèmes moteurs.!
•Il n’en est pas le cas: la phylogenèse a incrémenté le système moteur depuis la
périphérie jusqu’au niveau central.!
•Au cours de l’évolution, le système moteur a débuté par des protéines contractiles
seulement. Ces protéines contractiles se sont ensuite organisées en muscles, puis on a
sur 117

rajouté à ces effecteurs du système nerveux, des centres de commande. Sur ces
centres de commande ont été rajoutés d’autres centres de commandes, plus centraux
encore. !
•On est ainsi passé d’un système nerveux diffus à un système nerveux centralisé. !
•En suivant cette logique, il est facile de concevoir que même si le cerveau est le centre
commande le plus important, la moelle épinière reste capable de beaucoup de choses
car à un moment elle devait tout faire.!
•Logique du cours: muscle → cortex!
!
I - COMPOSITION DU SYSTÈME MOTEUR
•Le système moteur se divise en 4 sous-systèmes:!
-les circuits spinaux, faisant intervenir les interneurones et des groupes de
motoneurones!
-les systèmes descendants, faisant intervenir le cortex moteur et les centres du tronc
cérébral!
-les ganglions de la base, impliqués dans le filtrage des commandes appropriées du
mouvement!
-le cervelet, impliqué dans la coordination sensori-motrice.!
•La chaîne de commande du système moteur est composée des muscles, de la moelle
épinière, des centres du tronc cérébral et du cortex moteur. !
•Les ganglions de la base et le cervelet sont également impliqués dans la chaîne de
commande mais fonctionnent en parallèle.!
!
!A) La musculature!
!!1. Les différents types de fibres musculaires!
•La musculature se divise en:!
sur 217

-musculature lisse: Elle correspond à la musculature du système digestif et du
système cardiovasculaire.!
-musculature striée: C’est la musculature dite squelettique car elle est attachée au
squelette et permet dans la plupart des cas son mouvement (donc permet le
comportement). Il existe des exceptions: la plupart des muscles faciaux, notamment
la langue, sont des muscles squelettiques qui ne permettent pas le mouvement du
squelette. On peut distinguer la musculature striée en 3 grands types d’unités
motrices (= couples motoneurones-fibres musculaires):!
‣Unités motrices rapides et fatigables (FF: «$fast fatigable$»)!
‣Unités motrices lentes (S: «$slow$»)!
‣Unités motrices rapides et résistantes à la fatigue (FR: «$fatigue resistant$»)!
•Les muscles sont composés de ces différents types de fibres en proportion différente. La
proportion de ces différents types de fibres dépend du muscle concerné, mais est aussi
très variable d’un individu à l’autre (pour des raisons génétiques et en fonction de
l’activité de l’individu), et variable pour un même individu au cours du temps.!
•Ces trois types de fibres musculaires peuvent être différenciées selon la force
développée et leur capacité de maintenir cette puissance dans le temps.!
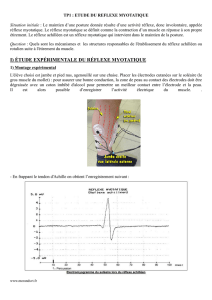
•La première figure exprime la force développée selon les différentes fibres, après une
stimulation unique:!
-Les fibres les plus puissantes sont les fibres rapides et fatigables.!
-Les fibres lentes sont les moins puissantes.!
-Les fibres rapides et résistantes développent une force intermédiaire.!
•Sur la deuxième figure, chaque vaguelette correspond à une stimulation. On voit à peu
près la même chose.!
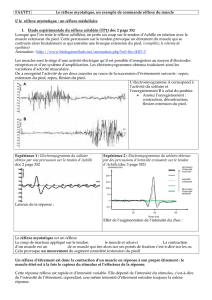
•La dernière figure exprime le pourcentage de la force maximale du muscle en fonction
du temps:!
-Les fibres rapides sont capable de conserver le maximum de leur puissance pendant
une quarantaine de secondes seulement. !
-Les fibres rapides et résistantes à la fatigue conservent une proportion raisonnable de
leur puissance maximale pendant un peu plus longtemps.!
-Les fibres lentes sont capables de conserver le maximum de leur puissance pendant
plusieurs heures.!
•Ces différences nous permettent de faire correspondre les types de fibres à certaines
activités: par exemple, les fibres rapides et fatigables correspondent à une activité très
explosive de type sprint. Les fibres lentes sont mobilisées pour des efforts qui se
sur 317

maintiennent dans le temps. En fonction de la longueur et de l’intensité de l’effort, les
muscles vont générer tel ou tel type d’unités motrices.!
!
!!2. Notion de synergie musculaire!
•La synergie est la capacité de coordonner les énergies. !
•Ce qui fait qu’on puisse obtenir le maximum de puissance au niveau d’un muscle, c’est
la capacité du système nerveux de correctement recruter les fibres musculaires
nécessaires à l’effort. !
•Cette synergie s’exprime:!
-au niveau du muscle, par le recrutement coordonné de plusieurs fibres musculaires
par un unique motoneurone!
-au niveau d’un groupe de plusieurs muscles, par la contraction coordonnée de
différents muscles destinée à exécuter un mouvement précis.!
•Note: Les activités musculaires qui donneront le rapport puissance/volume le moins
élevé correspondent au bodybuilding («$gonflette$»). Réciproquement, les activités
musculaires qui donneront le rapport puissance/volume le plus élevé sont les activités
de type escalade.!
!
!!3. La contraction musculaire!
sur 417

•La musculature striée est composée d’une suite de sarcomères séparés les uns des
autres par des stries Z (d’où le nom de muscle strié) avec deux types de filaments: des
filaments épais de myosine et des filaments fins d’actine. Lors de la contraction ces
filaments coulissent les uns par rapport aux autres.!
•L’arrivée du potentiel d’action au niveau du muscle provoque une libération de calcium.!
•Les filaments épais se finissent par des têtes de myosines. Ces têtes de myosine se
fixent et rampent sur les filaments d’actine. Ainsi, les sarcomères se raccourcissent et le
muscle aussi.!
•Les filaments d’actine sont munis de molécules de troponine capables de lier le calcium.
Suite à l’arrivée d’un potentiel d’action, la libération du calcium au niveau du muscle
contribue à la fixation des têtes de myosine sur les filaments d’actine.!
%
➤ Métabolisme énergétique impliqué!
sur 517
 6
7
8
9
10
11
12
13
14
15
16
17
6
7
8
9
10
11
12
13
14
15
16
17
1
/
17
100%