N°1 – Décrypter le vaisseau vivant

N°1 -
JANVIER 2005
Une publication
de la
Les mécanismes moléculaires sous-tendant la
formation des vaisseaux sanguins à caractère
segmentaire
Olivier Pourquié 3
La lymphangiogénèse au cours du
développement et de la maladie
Kari Alitalo et al. 4
Le rôle des récepteurs du guidage neural
dans le développement vasculaire
Anne Eichmann 6
Les cellules souches présentes dans les parois
des vaisseaux sanguins et impliquées dans
l'hématopoïèse et la régénération musculaire
Bruno Péault 8
L'utilisation de cellules souches liées aux
vaisseaux, les méso-angioblastes, dans la
réparation des muscles
Guilio Cossu 10
Les méso-angioblastes multipotents des
vaisseaux sanguins : l'origine des cellules
présentes dans l'aorte dorsale et le muscle
squelettique du somite
Margaret Buckingham et al. 11
Le rôle des cellules endothéliales
progénitrices dans la néovascularisation
Stefanie Dimmeler 13
Les cellules endothéliales progénitrices en
circulation : une source possible pour la
thérapie cellulaire autologue
Martine Aiach et al. 14
es maladies cardiovasculaires demeurent la première cause de
mortalité dans les pays occidentalisés et leur prévalence augmente
rapidement dans les pays moins développés. Malgré ce constat
alarmant, la morbidité et la mortalité cardiovasculaires ont nettement
diminué au cours des 40 à 50 dernières années : le nombre de
décès dus à des problèmes cardiovasculaires a chuté de près de 50 % grâce aux
progrès accomplis par la recherche cardiovasculaire dans plusieurs domaines, en
particulier dans la connaissance du système vasculaire. Il semble donc opportun de
consacrer une série de colloques à l'étude de l'arbre vasculaire, afin de dresser un
bilan des recherches en cours sur les étapes de la formation de l'arbre, les facteurs
impliqués dans celle-ci, la croissance coordonnée
de l'arbre, sa dégénérescence et sa fin, ainsi que
son éventuelle régénération au cours de la vie. Le
présent document offre un aperçu des débats du
premier de cette nouvelle série de colloques
Médecine et Recherche de la Fondation IPSEN,
consacré à l'origine et à la régénération de l'arbre
vasculaire et qui s'est tenu à Paris le 18 octobre
2004.
Les vaisseaux sanguins ont longtemps été
considérés comme des tubes passifs, soumis au
contrôle de stimuli neuraux ou d'hormones, jusqu'à
ce qu'on découvre qu'ils régulent leur propre
rythme en produisant localement des substances vasomotrices comme l'oxyde
nitrique (NO) dans l'endothélium ; cette découverte a marqué le début d'une
nouvelle ère en matière de physiologie et de pathologie vasculaires. La mise en
lumière du mécanisme intrinsèque de régulation de la paroi des vaisseaux a conduit
à reconnaître le dysfonctionnement endothélial comme une entité pathologique de
maladies dégénératives chroniques telles que l'hypertension, la dyslipidémie, le
Du développement
à la régénération
éditorial
sommaire
L
Décrypter le vaisseau vivant
Pierre Corvol
INSERM U36, Collège de France,
11, place Marcelin Berthelot,
75231 Paris, France
L’Arbre
vasculaire
Le nombre de décès dus
à des problèmes
cardiovasculaires a chuté
de près de 50 % grâce
aux progrès accomplis
par la recherche
cardiovasculaire dans
plusieurs domaines, en
particulier dans la
connaissance du
système vasculaire.
suite p.2

diabète de type 2 et l'athérome. L'athérome lui-même peut être considéré comme le prototype d'une
maladie complexe due à plusieurs facteurs, dans le cadre de laquelle se produit une inflammation de
la paroi vasculaire en réaction aux agressions biochimiques, toxiques et hémodynamiques.
L'interaction entre le développement des vaisseaux sanguins et les organes qu'ils irriguent est
complexe et inclut de possibles interactions réciproques avec le tissu adipeux et les nerfs en cours de
croissance. Les recherches menées au cours des 15 à 20 dernières années ont permis de faire la
lumière sur les premières phases du développement des vaisseaux lors de l'embryogenèse: le tube
vasculaire primitif est exclusivement composé de cellules endothéliales (vasculogénèse), avant de
former des artères, des veines et des capillaires à la suite de l'ajout de péricytes et de cellules
musculaires lisses vasculaires (angiogénèse), ou de devenir des vaisseaux lymphatiques. Plusieurs
gènes impliqués dans la vasculogénèse ainsi que dans la spécification des artères, veines et vaisseaux
lymphatiques ont été identifiés. Le rôle-clé assuré par les gènes codant pour les facteurs de
transcription nucléaires (protéines qui régulent l'expression d'un groupe d'autres gènes ayant des
fonctions liées) sensibles à la concentration en oxygène des cellules constitue un parfait exemple
d'interaction entre le tissu en formation et l'angiogénèse : les facteurs de transcription sont activés dans
les tissus hypoxiques et modifient l'expression de plusieurs gènes qui agissent de concert pour
déclencher le développement interne de nouveaux vaisseaux dans la zone privée d'oxygène. La
formation du système vasculaire subit également l'influence de forces hémodynamiques.
L'angiogénèse se produit également chez l'adulte. Il joue par exemple un rôle important à l'intérieur
de l'appareil reproducteur de la femme, dans la croissance et la différenciation du corps jaune, le
développement de l'endomètre pendant le cycle menstruel et la formation du placenta pendant la
grossesse. Il a aussi des implications pathologiques, puisque la croissance des tumeurs dépend en
grande partie de leur vascularisation, elle-même régulée par des facteurs de croissance vasculaire
produits par la tumeur. Les résultats obtenus sur plusieurs modèles expérimentaux confirment qu'un
traitement peut inhiber l'angiogénèse liée au cancer et à des maladies inflammatoires comme la
polyarthrite rhumatoïde ; par ailleurs, l'usage d'un anticorps monoclonal humanisé pour
lutter contre le facteur de croissance vasculaire endothélial (VEGF) a récemment été
approuvé pour les patients atteints d'un cancer du colon métastasé.
Autre progrès récent et porteur d'espoir : l'identification et la purification de
précurseurs (cellules souches et cellules progénitrices) de différents tissus composant le
système vasculaire adulte. L'existence de ces cellules vient étayer la théorie
controversée selon laquelle le système vasculaire, et en particulier le cœur, se
renouvellent continuellement tout au long de la vie. De plus, elle ouvre la perspective de
pouvoir utiliser ces cellules dans le cadre d'un traitement clinique pour favoriser la
régénération vasculaire chez des patients dont l'organisme n'est plus capable d'assurer lui-même ce
processus. Au cours des 10 dernières années par exemple, on a découvert des précurseurs
endothéliaux en circulation dans le plasma. Ils peuvent être isolés, cultivés ex vivo et, une fois injectés,
ils s'intègrent dans les zones où s'opère une néo-angiogénèse. Ces cellules font actuellement l'objet de
tests destinés à déterminer si elles peuvent favoriser le rétablissement de la circulation sanguine dans
des régions ischémiques ; certains des résultats obtenus sont aussi controversés que prometteurs [réf. 1,
2]. Plusieurs essais cliniques de stimulation de l'angiogénèse grâce à des gènes ou à des protéines
recombinées sont également en cours.
Ce compte-rendu de ce premier colloque reprend plusieurs de ces thèmes à travers trois parties. La
première concerne la formation du système vasculaire, et plus particulièrement le développement des
vaisseaux sanguins à caractère segmentaire (Olivier Pourquié), la lymphangiogénèse (Kari Alitalo) et
le guidage moléculaire de la croissance des vaisseaux sanguins (Anne Eichmann). La deuxième
partie décrit certains des types de cellules souches et de cellules progénitrices découvertes dans le
système vasculaire: les cellules souches issues des parois des vaisseaux sanguins et impliquées dans
l’hématopoïèse et la régénération musculaire (Bruno Péault) d’une part, et les cellules souches
associées aux vaisseaux, appelées méso-angioblastes (Guilio Cossu ; Margaret Buckingham) d’autre
part. Dans cette partie est également examinée la possibilité d’utiliser des cellules endothéliales
progénitrices dans le traitement de l’ischémie (Stefanie Dimmeler; Martine Aiach). La troisième section
aborde différents aspects du développement et de la régénération du cœur, au travers d’études sur la
formation de l’enveloppe cardiaque (Elisabetta Dejana) ainsi que sur les cellules souches
fonctionnelles, ou cardioblastes, présentes dans le cœur après la naissance (Kenneth Chien) et leur
potentiel d’aide à la régénération des cœurs lésés (Bernardo Nadal-Ginard). Un débat sur les aspects
pratiques de la thérapie cellulaire dans le traitement de l’insuffisance cardiaque (Philippe Menasché)
clôt cette partie. ◗
La ß-caténine, indispensable
au développement de
l'enveloppe cardiaque chez la
souris
Elisabetta Dejana et al. 16
Les cardioblastes post-nataux,
précurseurs de
cardiomyocytes entièrement
différenciés
Kenneth R. Chien et al. 18
L'auto-régénération du cœur :
les cellules souches
cardiaques et la régénération
du myocarde
Bernardo Nadal-Ginard et
Piero Anversa 20
La thérapie cellulaire dans le
traitement de l'insuffisance
cardiaque
Philippe Menasché 22
2L’Arbre Vasculaire - n°1 - Janvier 2005
Autre progrès récent et porteur
d'espoir : l'identification et la
purification de précurseurs (cellules
souches et cellules progénitrices) de
différents tissus composant le
système vasculaire adulte.
sommaire

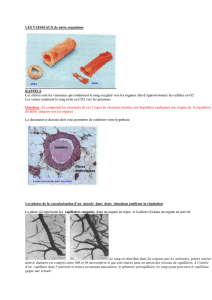
a répartition très caractéristique de
certains des principaux vaisseaux
sanguins reflète l'organisation de la
segmentation élémentaire de
l'embryon vertébré au début de son
développement. Ces segments, ou somites, sont
à l'origine de structures répétitives comme les
vertèbres et la musculature du tronc, ainsi que
des vaisseaux sanguins qui alimentent ces
structures. Après la gastrulation (c'est-à-dire
lorsque les trois feuillets embryonnaires,
l'ectoderme, le mésoderme et l'endoderme,
sont complètement formés), au fur et à mesure
de la croissance de l'embryon, les somites
commencent à former la partie dorsale du
mésoderme. Le mésoderme paraxial est
initialement indifférencié et la division cellulaire
se poursuit à l'intérieur, mais la partie située à
l'arrière de la région céphalique de l'embryon
évolue rapidement pour devenir le mésoderme
présomitique, à l'intérieur duquel se forment des
segments bien délimités et régulièrement
espacés. Ce processus progresse vers la région
caudale à un rythme régulier et sa durée
dépend de la taille du corps : 15 à 30 min par
segment chez le poisson zèbre, 90 min chez le
poulet, 120 min chez la souris et 4 à 5 h chez
l'homme. Une question essentielle se pose :
comment est induite la périodicité de ce
processus ?
Un gène marqueur spécifique du mésoderme
du poulet permet de constater qu'une vague
d'expression génique migre vers la partie
antérieure du mésoderme présomitique à
intervalles de 90 min [réf. 1]. Dans les années
1970, on expliquait la rythmicité de la formation
des segments à l'aide du modèle théorique
d'une horloge et de vagues ("clock-and-
wavefront") [réf. 2]. De récentes études sur
l'expression génique dans le mésoderme
présomitique confirment les suppositions de ce
modèle. L'horloge consiste en un rétrocontrôle
négatif entre deux voies de signalisation, Wnt et
Notch, dont l'activité cyclique et décalée forme
un oscillateur moléculaire [réf. 3]. La voie Notch
(un régulateur intracellulaire du développement
des cellules) favorise la maturation en
mésoderme paraxial, tandis que la voie Wnt
maintient les cellules du mésoderme
prismatique à l'état naïf (Fig. 1).
L'effet de vague en interaction avec l'horloge
a été prouvé grâce à des expériences de
microchirurgie menées sur des embryons de
poulet. L'inversion de petites parties du
mésoderme présomitique a démontré que
l'extrémité antérieure est vouée à se segmenter,
tandis que l'extrémité caudale reste plastique
[réf. 4]. Cette différence est due à un gradient
de la molécule, le facteur de croissance
fibroblasteique (FGF8), qui maintient les cellules
à l'état naïf. Bien que le gène fgf8 s'exprime sur
l'ensemble de ce gradient, la concentration de
la protéine du FGF8 est plus élevée dans les
cellules composant la partie caudale du
L’Arbre Vasculaire - n°1 - Janvier 2005 3
Les mécanismes moléculaires
de la formation des vaisseaux
sanguins
développement du système vasculaire
Olivier Pourquié
Stowers Institute for Medical Research.
1000 East 50th Street. Kansas City, Missouri, 64110,
Etats-Unis
L
Références
1. Palmeirim I, Henrique D, Ish-
Horowicz D, Pourquié O, Avian hairy
gene expression identifies a
molecular clock linked to vertebrate
segmentation and somitogenesis.
Cell 91, 639-648 (1997)
2. Cooke J, Zeeman EC, A clock and
wavefront model for control of the
number of repeated structures during
animal morphogenesis. Journal of
Theoretical Biology 58, 455-476
(1976)
3. Pourquie O, The segmentation
clock: converting embryonic time
into spatial pattern. Science 301,
328-330 (2003)
4. Dubrulle J, McGrew MJ, Pourquie
O, FGF signaling controls somite
boundary position and regulates
segmentation clock control of
spatiotemporal Hox gene activation.
Cell 106, 219-232 (2001)
5. Dubrulle J, Pourquie O, fgf8 mRNA
decay establishes a gradient that
couples axial elongation to
patterning in the vertebrate embryo.
Nature 427, 419-422 (2004)
L’analyse du développement embryonnaire montre l’importance
de certains gènes spécifiques, comme le fgf8, dans la croissance
et la structuration de l’arbre vasculaire.
MICRORÉSEAU
Technique destinée à analyser
simultanément l'expression d'un
grand nombre de gènes ;
fréquemment utilisée pour comparer
l'expression des gènes dans
différentes conditions (par exemple
l'âge, la pathologie).

mésoderme présomitique. Le gradient semble
résulter de la stabilité de l'ARN messager fgf8,
intermédiaire entre le gène
et la protéine ; or, cette
stabilité décroît dans la
partie antérieure du
mésoderme présomitique
[réf. 5], sans que l'on
connaisse la cause de cette
déstabilisation.
A un certain point sur le
gradient allant de la partie
caudale à la partie
antérieure du mésoderme,
le FGF8 atteint un seuil et
n'est plus suffisant pour
déclencher une réponse
cellulaire. Lorsque le signal
FGF8 est élevé, les cellules
restent à l'état naïf et ne peuvent pas réagir au
signal Notch périodique émis par l'horloge de
segmentation. Une fois que les cellules
atteignent la partie antérieure du mésoderme
présomitique, où le signal FGF8 est faible, elles
peuvent réagir au signal Notch et former un
nouveau domaine segmentaire, qui suit
immédiatement celui défini lors du précédent
cycle de l'horloge. La région segmentée
progresse donc vers la partie postérieure et le
processus recommence dans le mésoderme
présomitique juste derrière le segment qui vient
de se former. Chaque oscillation du cycle
Wnt–Notch, qui dure 90 min chez le poulet, crée
un segment. Nous cherchons à présent à
identifier d'autres gènes impliqués dans
l'oscillation, en comparant des microréseaux
d'ADN provenant d'une extrémité des embryons
à différentes phases du cycle de l'horloge et en
étudiant l'expression génique in situ à l'autre
extrémité de ces mêmes embryons. Une
dissection moléculaire aussi poussée devrait
bientôt permettre de décrire précisément la
régulation du processus définissant les
caractéristiques de base de nombreuses
structures, parmi lesquelles le système
vasculaire. ◗
4L’Arbre Vasculaire - n°1 - Janvier 2005
La lymphangiogenèse
au cours du développement
et de la maladie
développement du système vasculaire
Kari Alitalo et al.
Molecular/Cancer Biology Laboratory and Ludwig Institute for
Cancer Research, Haartman Institute and Biomedicum Helsinki,
P. O.B. 63 (Haartmaninkatu 8), 00014 University of Helsinki,
Finlande
e système lymphatique est un réseau
ouvert de vaisseaux transportant le
liquide interstitiel des tissus jusqu'aux
ganglions lymphatiques, avant de le
reverser dans le système vasculaire ; il joue un
rôle capital dans le drainage des tissus et la
surveillance immune. Il est également
largement impliqué dans la métastatisation des
tumeurs. Le décodage des mécanismes qui
régulent la formation du système lymphatique
chez l'embryon et la croissance des vaisseaux
lymphatiques chez l'adulte permettrait donc des
progrès conséquents dans le traitement des
œdèmes et l'arrêt de la propagation des
cellules cancéreuses.
Les vaisseaux lymphatiques sont recouverts
de cellules endothéliales espacées mais,
contrairement aux parois des vaisseaux
sanguins, ils ne comportent pas de vitrée, de
L
Figure 1 : Les voies de signalisation
Wnt et Nocht forment une horloge
moléculaire du développement.
GFP
protéine à fluorescence verte (green
fluorescent protein)
VEGF
facteur de croissance endothéliale
vasculaire (vascular endothelial
growth factor)
VEGFR
récepteur du facteur VEGF (types 1
à 3)
Toujours des
problèmes avec cette
image
Je dois voir cela avec
Astrid

L’Arbre Vasculaire - n°1 - Janvier 2005 5
lymphatiques présents dans la peau ont un
développement anormal, puisqu'ils ne sont pas
correctement reliés aux vaisseaux de drainage ;
cependant, un haut niveau de VEGF-C permet
de corriger cette anomalie chez la souris.
L'introduction du gène vegf-C dans les tissus
affectés grâce à un vecteur comme un
adénovirus ou un virus adéno-associé semble
être un traitement clinique prometteur. Une autre
anomalie héréditaire, la distichiasis
lymphœdème (LD), a une origine relativement
différente : une mutation du gène codant pour
le facteur de transcription FOXC2. Des études
menées sur des souris ont permis de constater
que cette voie empêche la production du
facteur de croissance dérivé des plaquettes, qui
attire les cellules musculaires lisses vers les
parois des vaisseaux [réf. 4]. Le gène foxc2
n'étant pas fonctionnel, les cellules
endothéliales des patients atteints de distichiasis
lymphœdème produisent le facteur de
croissance, ce qui entraîne une congestion des
petits vaisseaux lymphatiques par des cellules
musculaires lisses. On observe également chez
ces patients un problème lié aux valvules
présentes dans les vaisseaux lymphatiques et les
veines, mais on ignore comment la protéine
FOXC2 régule la formation de ces valvules.
Références
1. Saharinen P, Tammela T,
Karkkainen MJ, Alitalo K, Lymphatic
vasculature: development,
molecular regulation and role in
tumor metastasis and inflammation.
Trends in Immunology 25, 387-95
(2004)
2. Karkkainen MJ, Haiko P, Sainio K,
Partanen J, Taipale J, Petrova TV,
Jeltsch M, Jackson DG, Talikka M,
Rauvala H, Betsholtz C, Alitalo K.
Vascular endothelial growth factor C
is required for sprouting of the first
lymphatic vessels from embryonic
veins. Nature Immunology 5, 74-80
(2004)
3. Karkkainen MJ, Ferrell RE,
Lawrence EC, Kimak MA, Levinson
KL, McTigue MA, Alitalo K, Finegold
DN, Missense mutations interfere with
VEGFR-3 signalling in primary
lymphoedema. Nature Genetics 25,
153-159 (2000)
4. Petrova TV, Karpanen T, Norrmen
C, Mellor R, Tamakoshi T, Finegold D,
Ferrell R, Kerjaschki D, Mortimer P,
Yla-Herttuala S, Miura N, Alitalo K,
Defective valves and abnormal
mural cell recruitment underlie
lymphatic vascular failure in
lymphedema distichiasis. Nature
Medicine 10, 974-981 (2004)
Figure 2 : Mécanismes génétiques de l’angiogenèse et de sa métastatisation.
péricytes ni de cellules musculaires lisses. Les
cellules endothéliales des vaisseaux
lymphatiques et celles des vaisseaux sanguins
sont issues des mêmes précurseurs qui, chez
l'adulte, diffèrent par l'expression de quelque
200 gènes. Le développement des vaisseaux
lymphatiques et sanguins est régulé par
plusieurs membres étroitement liés de la famille
des molécules du facteur de croissance
endothéliale vasculaire (VEGF) [réf. 1]. Alors que
l'endothélium des vaisseaux sanguins réagit au
facteur VEGF classique via les récepteurs VEGFR-
1 et VEGFR-2 correspondants, les vaisseaux
lymphatiques, eux, contiennent presque
exclusivement le récepteur VEGFR-3, qui fixe les
facteurs VEFG-C et -D.
Le récepteur VEGFR-3 est présent dans les
angioblastes à un stade précoce du
développement du système vasculaire de
l'embryon et, à un stade plus avancé, dans
l'ensemble de l'endothélium ; il semble être
impliqué dans le remodelage du système
vasculaire de l'embryon aboutissant à une
hiérarchie de vaisseaux (angiogénèse). Au fur et
à mesure de la formation des vaisseaux
lymphatiques à partir des parois veineuses, les
cellules endothéliales commencent à produire
un facteur de transcription, le Prox-1, propre au
système lymphatique et qui accroît la réactivité
des cellules endothéliales au facteur VEGF-C. Au
même moment, le revêtement endothélial des
vaisseaux sanguins perd sa sensibilité au facteur
VEGF-C. A ce stade, la répartition du récepteur
VEGF-3 dans l'embryon suit de près le tracé des
vaisseaux lymphatiques et, chez les souris
n'ayant aucun des deux gènes pour le VEGF-3
ou pour son récepteur, les vaisseaux
lymphatiques ne se développent pas [réf. 2].
Toutefois, cela ne tient pas uniquement à la
présence ou à l'absence du facteur VEGF-C : les
souris possédant un gène vegf-c fonctionnel
survivent et ont un système vasculaire à peu
près normal, mais elles n'ont pas de vaisseaux
lymphatiques dans la peau, ce qui montre que
chaque tissu réagit différemment au niveau de
VEGF-C présent dans le corps. La peau semble
particulièrement sensible à ce niveau et les
vaisseaux lymphatiques y sont beaucoup plus
denses chez les souris stimulées afin de générer
dans la peau uniquement une concentration en
VEGF-C plus élevée.
Chez l'être humain, les lymphœdèmes, causés
par un drainage lymphatique insuffisant, sont
liés à un trouble héréditaire du développement
ou sont consécutifs à une intervention
chirurgicale ou à une maladie. Le
lymphœdème primaire (syndrome de Milroy)
est attribué à des mutations du gène vegfr-3, qui
réduisent l'efficacité du VEGFR-3, le récepteur du
facteur VEGF-C [réf. 3]. Les vaisseaux
Plus de 200 gènes régulent le cycle de vie des cellules
endothéliales des vaisseaux sympathiques. La famille moléculaire
des facteurs de croissance endothéliale vasculaire (VEGF) joue
notamment un rôle clé. La maîtrise de ces mécanismes
moléculaires ouvre un vaste pontentiel thérapeutique.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%