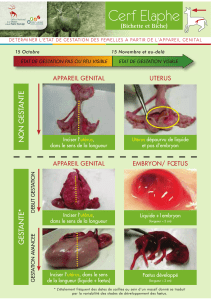
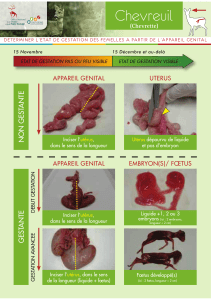
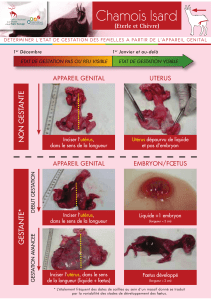
II. Deuxième partie : Description des différents types de gestation

VETAGRO SUP
CAMPUS VETERINAIRE DE LYON
Année 2014 - Thèse n°
LA GESTATION EXTRA-UTERINE CHEZ LES MAMMIFERES
DOMESTIQUES ET LA FEMME
THESE
Présentée à l’UNIVERSITE CLAUDE-BERNARD - LYON I
(Médecine - Pharmacie)
et soutenue publiquement le 9 juillet 2014
pour obtenir le grade de Docteur Vétérinaire
par
WALLER Laureline
Née le 22 mai 1990
à Valréas

2

3

4

5
REMERCIEMENTS
A Monsieur le Professeur Jean-François GUERIN
De la Faculté de Médecine
Qui nous a fait l’honneur d’accepter la présidence de notre jury de thèse
Qu’il trouve ici l’expression de ma reconnaissance ainsi que de mes sincères remerciements
A Monsieur le Professeur Pierre GUERIN
Du Campus Vétérinaire de Vetagro Sup
Qui nous a fait l’honneur d’encadrer ce travail
Pour son soutien, sa disponibilité, sa gentillesse et ses conseils avisés depuis le début de ce projet
Qu’il trouve ici l’expression de mes remerciements chaleureux et de mon plus grand respect
A Monsieur le Docteur Pierre BRUYERE
Du Campus Vétérinaire de Vetagro Sup
Qui nous a fait l’honneur de participer à notre jury de thèse
Pour m’avoir aidé, encouragé et accepté de juger ce travail avec justesse
Qu’il trouve ici l’expression de mes remerciements les plus sincères
Au Docteur Emilie ROSSET
Pour son aide précieuse et son autorisation à l’utilisation d’un de ses cas cliniques. Sincères
remerciements.
Au Docteur Delphine GREZEL
Pour son implication, ses conseils avisés et son aide sur ce travail. Sincères remerciements.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
1
/
88
100%