p2-u4-gazaille-td1-physiologie-efr-18-11-16-word

1
UE4 – Appareil respiratoire
GAZAILLE
Date : 18/11/16 Plage horaire : 14H - 15h30
Promo : DFSGSM2 2016/2017
Enseignant : Dr GAZAILLE
Ronéistes :
GRUAU Antoine
LE BARS Coralie
TD : Physiologie EFR (partie 1)
I. Rappel
1.1 Structure de l'appareil respiratoire
1.2. Les voies aériennes
1.3. Les vaisseaux sanguins
1.4. L'interface gaz sang
II. Volumes pulmonaires et ventilation (cf EFR)
2.1. Technique de mesure
2.2. Ventilation = renouvellement des gaz pulmonaires
2.3. Gaz alveolaire et volume mort physiologique
2.4. Ventilation alveolaire v'a
2.5. Pression partielle des gaz alveolaires
A. Gaz carbonique
B. Oxygène
C. Équation des gaz alvéolaires
III. Circulation pulmonaire
3.1. Pression, debit et resistances dans la circulation pulmonaire

2
3.2. Pression gazeuse pulmonaire
3.3. Gravite
3.4. Vasomotricite
IV. Diffusion alvéolo capillaire
4.1. Diffusion de l’O2
4.2. Diffusion du CO2
V. Rapports ventilation/perfusion
5.1. Rappel
5.2. Effets des modifications du rapport ventilation/perfusion
A. Transport de l'Oxygène
B. Transport du dioxyde de carbone
VI. Mécanique ventilatoire
6.1. Maintien et mobilisation des poumons
6.2. Muscles inspiratoires
A. Autres muscles inspiratoires
B. Diaphragme
6.3. Muscles expiratoires
6.4. Relation entre pression et volumes : notion de compliance
I. Rappel
1.1. Structure de l'appareil respiratoire
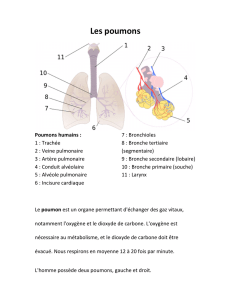
La respiration (=transfert de gaz/procurer de l'O2/éliminer le CO2) comprend la respiration
cellulaire (cf cycle de Krebs et mitochondries, ne reviendra pas dessus), les échanges gazeux
(diffusion du CO2 de l'organisme vers l'extérieur et de l'O2 de l'extérieur vers l'intérieur) et la
respiration externe ou ventilation.
On part de quelque chose qui est macroscopique (flux d'air entre l’extérieur et l'intérieur de
l'organisme) puis échelle de l'organe (transfert de gaz) et enfin niveau cellulaire (consommation
d'oxygène).

3
(Nb : 3 bronches lobaires droites vs 2 bronches lobaires gauches)
1.2. Les voies aériennes
Les voies aériennes (VA) commencent à partir du nez ou de la bouche, jusqu'aux alvéoles :
_ VA supérieures : au dessus de la glotte
_ VA inférieures : trachée jusqu'aux bronchioles terminales
Le volume mort anatomique (l'air aspiré qui ne participe pas aux échanges physiologiques) est
d'environ150mL
_ Bronchioles respiratoires (canaux alvéolaires bordés d'alvéoles, avec en aval l'unité anatomique
=acinus)

4
Tableau : plus on descend dans l'arbre bronchique, plus le calibre se réduit et plus la surface de
section (en cm²) augmente.
La multiplication des subdivisions fait qu'au final on a, au niveau des sacs alvéolaires, une surface
de 800 000 cm².

5
Bronches avec plaques cartilagineuses (bleu)
1.3. Les vaisseaux sanguins
Les vaisseaux sanguins se ramifient depuis l'artère pulmonaire jusqu'au capillaire se drainant dans
les veinules pulmonaires.
Anatomiquement :
_ Les artères suivent les ramifications bronchiques
_ Les veinules et veines cheminent dans la paroi des lobules
_ Les capillaires forment un réseau dense (diamètre de 10 microns)
1.4. L'interface gaz sang
Les échanges gazeux se font au niveau du sac alvéolaire. O2 et CO2 se déplacent entre le gaz
alvéolaire contenu dans l'acinus et le sang contenu dans les capillaires.
L'épaisseur de la membrane alévolo-capillaire < 1micron et surface importante : 80 à 100m² .
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
1
/
22
100%