Métabolisme carboné

Métabolisme carboné
I] La biosynthèse des glucides
Une fois que les triosephosphate sont sortis du chloroplaste, ils vont engendrer le pool des hexose :
2 triosephosphates vont se condenser en fructose 1,6-bisphosphate, qui va se déphosphoryler, et va se convertir réversiblement
en glucose-6-phosphate. A partir de ce pool, une grande partie des métabolismes végétaux vont s'établir.
1) La synthèse du saccharose
La majeur partie du carbone fixé se retrouvera dans le saccharose.
C'est la molécule de « transport à longue distance », synthétisé dans
la feuille, il ira alimenter les parties non photosynthétiques. Ce n'est
pas un glucide réducteur, la fonction aldéhyde étant impliqué dans
la liaison glycosidique. Certaines plantes stockent de grandes
quantités de saccharose (canne à sucre, betterave, carotte...).
La synthèse du saccharose implique l'UDP-Glucose dans deux voies
métaboliques :
•L'UDP-Glucose est condensé au fructose-6-phosphate par
la saccharose-phosphate synthase pour faire du saccharose
phosphate, et le saccharose-phosphate phosphatase amène
à la synthèse de saccharose. La saccharose phosphate
synthase est activé par le glucose-6-phosphate et une
déphosphorylation, et inhibé par une phosphorylation
(régulation post-traductionnelle).
UDP−Glucosefructose−6−phosphate
saccharose−phophate synthase saccharose−phosphate
saccharose−phosphate
saccharose−phosphate phosphatase saccharose
•UDP-Glucose et fructose sont condensés par Susy (sucrose-synthase) pour donner saccharose + UDP. Cette réaction
est réversible.
Toutes ces réactions sont exergoniques, donc le bilan énergétique est positif de 25kJ/mol, donc thermodynamiquement quasi
irréversible. La synthèse d'UDP-galactose se fait également, et l'UTP nécessaire à la formation d'UDP-Glucose est régénéré en
consommant de l'ATP¨:
UDPATP UTPADP
2) La synthèse de l'amidon dans les amyloplastes
Dans le cas de la synthèse d'amidon transitoire, les
produits d'origines étaient faits sur place. Dans ce cas là,
c'est les glucides de la sève élaborée qui seront utilisés,
avec nécessité d'un transporteur à la surface des
amyloplastes : il s'agit d'un antiport glucose-6-
phosphate/Pi. Pour éviter une augmentation de la pression
osmotique, les amyloplastes polymérise les glucoses en
amidon :

glucose−6−phosphate
isomérase glucose−1−phosphate
glucose−1−phosphate
ADP −glucose pyrophosphorylase ADP−glucose amidon
On remarque que le glucose-6-phosphate est également
le substrat de la formation du glycogène chez les
bactéries, renforcant la théorie endosymbiotique de la
genèse des plastes. Chez les végétaux, on a deux formes
de stockage : l'amylose (α 1,4-glucose) et amylopectine
(amylose + α 1,6-glucose).
Deux systèmes enzymatiques permettent la formation
d'amidon, une enzyme soluble (majoritaire), et une
enzyme granulaire qui se fixe sur le granule en
formation. Des enzymes de branchement permettraient
d'obtenir des amylopectines peu branchées (assez lent),
et des amylopectines très branchées (rapidement).
Actuellement, on pense que le système de branchement
effectue ces modifications lentement.
3) La migration des assimilats
Les assimilats migrent depuis les organes sources vers les organes puits par la sève
élaborée, au travers du phloème constitué de cellules criblées qui ont une différenciation
particulière : le noyau se détruit, les structures cytoplasmiques se modifient
considérablement (rétraction puis disparition de la vacuole due à la dégénérescence du
tonoplaste), les parois transversales vont être partiellement dégradée (crible) pour mettre une
jonction plus efficace entre les différentes cellules. La membrane elle reste intacte, mettant
les cytoplasmes en communication les uns avec les autres. Dans ces cellules criblées, des
protéines s'accumulent, ayant pour rôle de cicatriser les tissus conducteur et défendre.
Les cellules criblées sont accompagnées de cellules compagnes avec un cytoplasme très
dense. Ces deux cellules sont issues d'une même cellule mère. La plante, dans certaines
conditions, est capable de fermer les cribles avec de la callose (lors des saisons froides par
exemple). Le saccharose passe dans les cellules compagnes (puis les tissus) par la voie du
symplasme, le reste du transport par la voie de l'apoplasme.
Le xylème est lui issu de la mort programmé, avec lignification des parois.
L'eau passe du phloème au xylème au bas de la plante près des organes cibles (car peu de
POA), et inversement près des organes source (car beaucoup de POA). La descente du
saccharose se fait par la pression hydrostatique. Dans le phloème, c'est une circulation sous
pression, et dans le xylème, une circulation sous tension (évapotranspiration).
4) Les navettes plaste/cytosol
Les chloroplastes vont pouvoir fournir de l'ATP au
cytoplasme, avec des protéines sur la membrane
interne du plaste capable d'échanger ATP/ADP.
NADPH incapable de traverser la membrane du
chloroplaste. On a plusieurs navettes :
Oxaloacétate/malate : d'une part
consomme du pouvoir réducteur dans le
chloroplaste et en génère dans le
cytoplasme
Aspartate-cg/glu-OAA + MAL/CG :
transport couplé permettant aussi la
génération de pouvoir réducteur dans le
cytosol.
Au niveau de l'amyloplaste, ces
transports permettent l'importation de

pouvoir réducteur et d'ATP nécessaires à la synthèse d'amidon.
II] Le métabolisme glucidique
Une grande partie des molécules glucidiques vont être utilisés comme substrats respiratoire.
1) La dégradation du saccharose et de l'amidon
Lorsque la plante va devoir récupérer ces molécules, elle va en
extraire les monomères du glucose. Le saccharose peut être
dégradé de deux manières :
•Susy : action réversible lorsque la plante a besoin de
glucose
Invertase (pH 7,4 pour les cytosolique, pH acide pour les
pariétales et plastidiales) : coupe le saccharose en ses deux
monomères. C'est irréversible.
A l'heure actuelle, on ne sait pas vraiment si l'un
de deux systèmes est favorisé...
La dégradation de l'amidon doit amener du
glucose :
•Sur l'amylose : α-amylase coupe au
milieu l'amylose en dextrines jusqu'au
maltose et l'α-glucosidase en fera du
glucose. La β-amylase coupe dextrine
et amylose en bout de molécule
directement en maltose. Ce système
permet d'optimiser la dégradation en
augmentant les possibilités de fixation
des β-amylase.
•Sur l'amylopectine : α et β-amylase
coupent en dextrines branchés, des
enzymes de débranchement
débranchent pour faire des dextrines,
qui seront coupées par α et β-amylase.
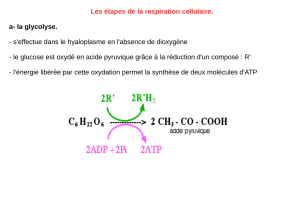
2) La glycolyse
Le glucose est phosphorylé par un ATP,
puis isomérisé en fructose-6-phosphate.
Il est phosphorylé en fructose-1,6-
bisphosphate. Il donne DHOAP et
AldPG. DHOAP passe en AldPG. Il
passe en BPGA puis en 3-PGA avec
synthèse d'ATP. On passe ensuite au 2-
PGA puis au PEP et enfin au pyruvate
avec synthèse d'ATP. Le PEP pourra
alors entrer dans le cycle de Krebs.
Phosphorylation liée au substrat :
possibilité pour les cellules de générer
de l'ATP sans passer par les ATP
synthase.
La respiration est une réaction de
catabolisme, entraînant une dégradation
complète du substrat en composés
uniquement minéraux, et où l'accepteur
terminal des électrons transférés au

cours des oxydo-réductions est l'oxygène.
3) Les caractéristiques de la respiration
On peut parler du quotient de respiration :
QR=volume de CO2dégagé
volume de CO2absorbé
On a deux types de respiration selon le stade de développement. Globalement, l'intensité respiratoire va dépendre de l'âge des
organes et des conditions de l'environnement. La respiration sera très faible dans les graine mais augmenter très fortement
durant la germination. Elle sera généralement très élevée dans les organes en croissance, et particulièrement les méristèmes. On
va donc considérer deux respiration : la respiration de croissance (pour 1g de protéine, on a besoin de 2,5g de glucose et de 3
gramme pour les lipides...) et la respiration de maintenance (fournit de l'énergie nécessaire à l'entretien des fonctions de la
plante). Ainsi, d'un tissus à un autre, l'intensité respiratoire peut beaucoup varier. Les échanges respiratoires se font plus
lentement que les échanges de gaz lors de la photosynthèse (d'un rapport 10).
4) Le cycle de Krebs
Cette réaction se fait dans la mitochondrie. On condense le PEP en
Acetyl-CoA (avec premier dégagement de CO2). On passe au citrate,
ensuite une isomérisation par l'aconitase pour donner l'isocitrate. Une
deuxième décarboxylation et formation de NADH pour donner un
kétoglutarate. On a a nouveau adjonction de CoA, avec une nouvelle
décarboxylation et formation d'une troisième molécule de NADH pour
donner le succinyl CoA. Par phosphorylation on passe au succinate
avec dégagement de GTP. On passe au fumarate avec dégagement de
FADH2, au malate et à l'oxaloacétate avec dégagement de NADH.
Pour une molécule de glucose, on a 8 NADH, 2 FADH2 et au niveau
de l'ATP 2 sont générées par la glycolyse et 2 par cycle de Krebs.
Les réactions anaplérotiques du cycle de Krebs : à partir de la PEP
dans les plantes, on peut avoir de l'OAA (grâce à la PEPc) puis du
malate, chacun de ces composés pouvant être injecté dansa le cycle de
Krebs.
5) La chaine de transfert des électrons et la synthèse d'ATP
Le complexe I (NADH ubiquinone
réductase) utilise une ubiquinone pour
faire sortir des protons.
Le complexe II fait entrer du FADH2.
Le complexe III (ubiquinone
cytochrome c réductase) avec un cycle
semblable au cycle Q.
Le complexe IV est une cytochrome c
oxydase, composée de deux molécules
de cytochrome (3 et a3) : c'est
l'oxydase terminale.
Le complexe V est une ATP synthase
qui utilise le gradient de proton pour
former de l'ATP.
La rotenone est inhibitrice du
complexe I (inhibitrice des complexes
NADH dépendants). Le cyanure et
l'azide inhibent le complexe IV.
Dans les cellules végétales, il y a deux systèmes de type NADH déshydrogénase supplémentaire, un sur la face interne, l'autre
sur la face externe (qui pourra travailler avec les molécules de NADH venant du cytoplasme, et de la glycolyse notamment).

Ces systèmes permettent entre autre de contourner l'inhibition par la roténone.
Le complexe I et le complexe III transfèrent 4 protons par NADH, et le complexe IV en injecte 2. Chaque NADH injectera 10
protons dans l'espace inter-membranaire. Il faut 3 proton par ATP, ainsi, un NADH formera 3 ATP. A chaque fois qu'un FADH 2
entre, on aura plus que 6 protons, donc 2 ATP. Il en est de même pour les NADH déshydrogénase de la face externe.
Si on fait le bilan énergétique :
•2 ATP par le cycle de Krebs,
•2 ATP par la glycolyse
•24 ATP par les 8 NADH synthétisés par le cycle de Krebs
•4 ATP par les deux FADH2.
•4 ATP par les NADH de la glycolyse (pris en compte par la déshydrogénase externe)
•
On a bien 36 molécules d'ATP pour une molécule de glucose chez les végétaux, soit 1050 kJ. Le rendement est de 37 %
(oxydation totale du glucose à 2870 kJ).
Les mitochondries végétales sont capables de respirer en présence du cyanure, grâce à une oxydase alternative capable de
récupérer les électrons de l'ubiquinone, et de les transférer directement à la molécule d'oxygène, court-circuitant complètement
le complexe III et le complexe IV. Cependant, ce système est peu générateur d'énergie (seulement par le complexe I). Cette
protéine est un dimère, chaque monomère contenant un atome de fer (responsable du transfert d'électron), capable de
reconnaître les ubiquinones et l'O2 (affinité 10 fois inférieur que Complexe IV). Les monomères peuvent être liées par un pont
disulfure (peu active) ou réduite (très active). On considère actuellement que la réduction de ce pont disulfure est une manière
de réguler ce système. Chez l'Arum, la thermogenèse des fleurs (rôle pollinisateur) utilise l'oxydase, limitant la genèse d'ATP
mais permettant la dissipation de l'énergie des composés réduits non utilisés.
6) Les navettes mitochondriales
La plupart de l'ATP synthétisée va être exporté vers le
cytosol par un antiport ATP/ADP. Des transporteurs de
NADH permet l'entrée du NADH pour éviter de court-
circuiter le complexe I (bien que ce processus soit
minoritaire), une par le biais du Malate/oxaloacétate
(équilibre du pouvoir réducteur de chaque côté) qui permet de
transférer le pouvoir réducteur, et une glutamate/aspartate.
7) La régulation de la respiration dans les mitochondries végétales
La vitesse de la respiration mitochondriale va être contrôlé par la charge énergétique cellulaire (ATP/ADP). Ainsi, dans une
cellule métaboliquement très active, la respiration sera forte. Au contraire, elle sera très faible dans les cellules quiescentes.
D'un côté, la voie des cytochromes génère beaucoup d'ATP, et la voie alternative moins.
On peut donc penser que cette voie alternative à d'autres objectifs :
•La cellule a nécessité de maintenir un équilibre redox : accumulation de pouvoir réducteur sans nécessité de synthèse
d'ATP. La voie alternative peut donc réguler le pouvoir réducteur, sans déséquilibrer la charge énergétique. Dans la
maturation des fruits, on trouve deux types : les climactériques (forte augmentation de leur intensité respiratoire lors
d'un stade de la maturation allant de paire avec l'éthylène) utilisent essentiellement l'oxydase alternative.
•Cette oxydase alternative a également un rôle dans la réponse au stress : dans les mitochondries, des formes actives de
l'oxygène sont générées. Or on sait maintenant que ces espèces ont un rôle dans la signalisation cellulaire. La voie
alternative baisse les chances de générer des espèces actives de l'oxygène. Ainsi, cette voie permet donc de réguler la
réponse au stress des plantes.
8) Les fermentations alcooliques et lactiques
Dans certaines conditions, les plantes devront passer d'un métabolisme aérobie à anaérobie, situation devant être transitoire. Le
riz va germer en anaérobiose, nécessitant un métabolisme de fermentation pour la croissance de la première feuille et du
 6
7
6
7
1
/
7
100%