File

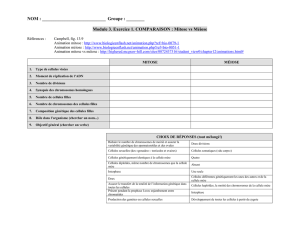
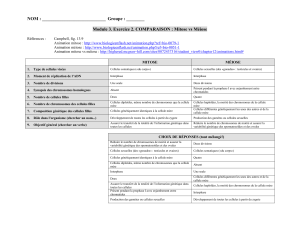
1 cellule mère (2n)
La méiose
Entrecroisement
(
crossing
-
over)
Réplication
de l’ADN
2n chromosomes simples
2n chromosomes doubles
4 cellules filles (1n) 1n chromosomes simples
(
crossing
-
over)
Méiose 1
Méiose 2
1n chromosomes doubles

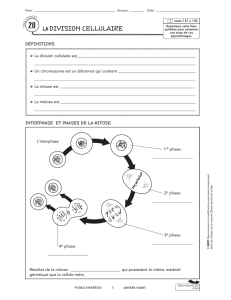
Les phases de la méiose
M1 : Division réductionnelle
Prophase I
Métaphase I
Anaphase I
Interphase II
Télophase I

Les phases de la méiose
M2 : Division équationnelle
Prophase II
Métaphase II
Anaphase II
Télophase II
Interphase III


 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
1
/
29
100%