viii-1. electrophysiologie veterinaire - Lodel

Œil et Physiologie de la Vision - VIII-1
VIII-1. ELECTROPHYSIOLOGIE VETERINAIRE
Serge Georges Rosolen
Pour citer ce document
Serge Georges Rosolen, «VIII-1 : ELECTROPHYSIOLOGIE VETERINAIRE», Oeil et
physiologie de la vision [En ligne], VIII-La vision et son exploration chez l'animal, mis à
jour le 18/06/2013, URL :
http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=213,
doi:10.4267/oeiletphysiologiedelavision.213
Plan
Introduction
Intérêt de l’ERG en médecine vétérinaire
But du chapitre
Physiologie animale comparée
La rétine
Généralités
Quelques particularités des rétines animales
Les voies visuelles
Description anatomofonctionnelle
Exploration rétinienne par ERG
Les conditions de genèse de l’ERG flash
Le choix du stimulateur et de la stimulation
Les conditions de stimulation
Les conditions de recueil et de traitement du signal
Les protocoles
Tester la réponse du système photopique
Tester la réponse du système scotopique
Tester la réponse des deux systèmes
Tester selon les espèces
Les facteurs affectant l’ERG flash
Les composantes de l’ERG et leurs origines
Indications de l’ERG flash chez l’animal
Exemples de résultats normaux
Situations cliniques
Le fond d’œil n’est pas visible
Les affections rétiniennes hérédodégénératives
Les anomalies génétiques des affections rétiniennes
Les troubles du comportement
Autres tests électrophysiologiques
Les autres ERGs
Les PEV
Contraintes anatomiques
1

Œil et Physiologie de la Vision - VIII-1
Contraintes techniques
Conclusion
Texte intégral
Remerciements au Professeur Jean Sautet pour sa contribution aux illustrations d'anatomie
Introduction
L’appréciation de la fonction visuelle chez l’animal se fait sur des critères subjectifs
complétés par des tests subjectifs et objectifs.
Les critères subjectifs sont variés. On peut observer par exemple l’isolement d’un
individu au sein d’un troupeau de bovins ou d’ovins, l’apparition soudaine d’une
agressivité à l’encontre d’autres individus, une diminution notable de l’activité de jeu ou
de travail comme pour le chien de chasse ou encore une perte de poids chez un individu
au sein d’une colonie en élevage comme pour les rats, les souris ou les singes…
Ces signes déclenchent la mise en œuvre de tests subjectifs individuels d’évaluation de la
fonction visuelle comme la recherche du réflexe de clignement à la menace ou le test de
la boule de papier en vision binoculaire. Ces tests subjectifs individuels peuvent être
entrepris chez des espèces comme le chien ou le chat mais pas chez les espèces « mal
voyantes » comme les rongeurs (rat, souris, …). Dans le cas de ces espèces, importantes
pour la recherche biomédicale, il est nécessaire d’évaluer le fonctionnement de leur
système visuel par des tests objectifs électrophysiologiques.
Certains de ces tests électrophysiologiques sont utilisés en ophtalmologie vétérinaire,
c'est-à-dire pour les animaux de compagnie (chien, chat, cochons d’inde, lapins, etc.), de
rente (moutons, bovins, etc.) et chez certaines espèces sauvages maintenues en
captivité (oiseaux, primates, félins, etc.).
Ils permettent une évaluation objective essentiellement la fonction rétinienne de l’animal,
à l’aide de l’enregistrement de l’électrorétinogramme (ERG).
Intérêt de l’ERG en médecine vétérinaire
Les progrès de la médecine et de la chirurgie vétérinaires notamment en d’ophtalmologie
(chirurgie de la cataracte par incision étroite, mise en place d’implants intraoculaires,
etc) d’une part et la prise en compte, par la société, de l’animal en tant qu’être vivant
d’autre part, font que celui-ci est considéré comme un véritable patient, pouvant
bénéficier des mêmes approches diagnostiques et des mêmes stratégies thérapeutiques
que celles utilisées pour l’espèce humaine.
L’ERG est un examen complémentaire s’inscrivant dans cette démarche d’unicité de la
médecine [Zinsstag et al., 2005], [Cardiff et al., 2008] pour un bénéfice réciproque des
traitements de l’homme et de l’animal considéré comme un patient à part entière
[Enserink, 2007], [Rosolen SG, Picaud, 2010].
2

Œil et Physiologie de la Vision - VIII-1
But du chapitre
Ce chapitre a pour but de fournir les éléments essentiels à l’enregistrement et à
l’interprétation des examens électrorétinographiques effectués chez différentes espèces
d’animaux de compagnie atteints de maladies homologues ou non à des affections
rencontrées dans l’espèce humaine essentiellement des chiens et des chats.
L’enregistrement d’ERG effectués dans des conditions standards de stimulation, de
recueil et de traitement du signal est souvent indispensable pour le diagnostic et le
pronostic des affections concernées.
Au préalable, quelques caractéristiques anatomofonctionnelles visuelles animales sont
présentées, ainsi que les conditions spécifiques de mise en œuvre et de réalisation des
examens électrorétinographiques.
En fin de chapitre, quelques exemples de résultats sont commentés, enregistrés
essentiellement chez le chien.
Physiologie animale comparée
La rétine
Généralités
Bien que l’éventail des perceptions sensorielles des animaux soit très riche, c’est
néanmoins avec notre cerveau d’homme et non de poisson, d’oiseau, de chien ou de
cheval que nous analysons la façon dont les animaux se servent des informations reçues
et les interprètent.
Nous devons donc être très prudents dans l’interprétation des résultats des examens
pour l’évaluation de la fonction visuelle des espèces animales comme les vertébrés.
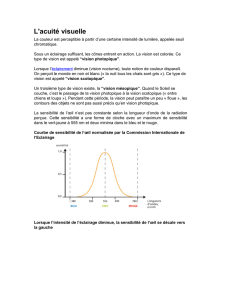
L’œil dans l’évolution
Au cours de l’évolution, l’œil et la fonction visuelle se sont avant tout adaptés au milieu
aquatique, terrestre ou aérien et au mode de vie nocturne ou diurne, de chaque espèce.
L’œil de type camérulaire
Chez tous les vertébrés, l’œil est de type camérulaire : c’est un objectif (le segment
antérieur avec ses lentilles et son diaphragme) et une chambre noire (le segment
postérieur) comportant un écran protecteur (absorbant l’excès de photons incidents) et
un capteur de photons (la rétine) qui transforme l’information lumineuse (énergie et
longueur d’onde) en un signal électrophysiologique (influx nerveux) interprétable par le
cerveau.
Les figures VIII-1, VIII-2, VIII-3 montrent les différents types d’yeux des vertébrés
aboutissants, chez les mammifères, au type camérulaire.
Figure VIII-1. L’œil de poisson (a) et (b) est gros et constitué d’une coque sclérale très
développée chez les espèces abyssales, permettant une résistance à la pression. Il
caractérisé par une cornée aplatie et un volumineux cristallin de forme sphérique non
déformable (a). Comme le cristallin n’est pas déformable, la mise au point
(accommodation) ne peut se faire que grâce à son déplacement d’avant en arrière à
l’aide d’un muscle (b).
3

Œil et Physiologie de la Vision - VIII-1
Lors de la conquête du milieu aérien par les amphibiens (c) et les reptiles (d), les glandes
lacrymales et les paupières apparaissent. Elles permettent une hydratation de la cornée.
La cornée se bombe et cette nouvelle conformation (modification du rayon de courbure)
permet de s’adapter au changement d’indice de réfraction entre l’extérieur (air) et
l’intérieur (aqueux)… L’accommodation se fait par déplacement du cristallin, comme pour
les poissons.
Figure VIII-2. Chez les mammifères (e), le cristallin peut se déformer grâce à un système
complexe de muscles. Le mode de vie conditionne la forme et le volume du cristallin.
Chez les mammifères diurnes (f), le cristallin est lenticulaire alors que chez les espèces
nocturnes (g) (lémurien, rongeurs), le cristallin est sphérique et très volumineux (g) avec
une pupille en mydriase permanente qui laisse entrer une quantité maximale de lumière.
Figure VIII-3. Le système le plus élaboré se rencontre chez les oiseaux plongeur/pêcheur
comme le cormoran (h). L’œil est très volumineux, il occupe tout l’orbite. Lorsque
l’oiseau est en plongée, le muscle de Crampton permet l’aplatissement de la cornée
comme pour les poissons. Sous l’action du muscle de Brüch, le cristallin peut se déformer
jusqu’à former un lenticône. Le cormoran peut ainsi faire varier la puissance optique de
ses yeux d’environ 50 dioptries.
Le tableau VIII-1 résume les différents éléments anatomiques de l’œil des différents
vertébrés expliquant leur adaptation au milieu et leur mode de vie.
L’œil mobile
La vision n’est efficace que s’il y a mouvement car les capteurs visuels ne réagissent
qu’aux changements de luminosité. Ce mouvement peut être celui de l’objet observé ou
celui de l’œil lorsque l’objet est immobile. Dans ce cas, il y a déplacement de l’image sur
la rétine par des saccades oculaires.
Quelques particularités des rétines animales
La rétine inversée des vertébrés
Chez tous les vertébrés, la rétine, présente à quelques variations près, le même schéma
fonctionnel (figure VIII-4) : a) un étage préréceptoral, b) un étage réceptoral composé
d’une très fine mosaïque de deux types de photorécepteurs : cônes et bâtonnets, c) un
étage postréceptoral avec cellules horizontales, bipolaires et amacrines et d) un étage de
conduction du message à partir des cellules ganglionnaires jusqu’au cortex visuel
primaire puis secondaire, très développé chez les mammifères.
La densité des deux types de photorécepteurs varie en fonction des espèces. Le tableau
VIII-2 présente ces différentes densités selon les différentes espèces et les auteurs
[Steinberg et al., 1973], [Wikler et al., 1990], [Zhang, Straznicky, 1991], [Jonas et al.,
1992], [Famiglietti, Sharpe, 1995], [Chandler et al., 1999], [Mowat et al., 2008].
Fovéa et area centralis
Animaux diurnes
La rétine des animaux diurnes (poissons, reptiles, oiseaux, carnivores, herbivores,
primates) est équipée de cônes nombreux, denses et diversifiés leur conférant la
perception des couleurs et une bonne acuité visuelle [Ebrey, Koutalos, 2001], [Wright,
Bowmaker, 2001] : poissons et oiseaux sont trichromates et la plupart des mammifères
sont dichromates (sauf l’homme et certains primates) [Packer et al., 1989], [Curcio et
al., 1990], [Jacobs, 1993], [Szel et al., 1996], [Ahnelt, Kolb, 2000].
4

Œil et Physiologie de la Vision - VIII-1
Une voire deux fovéas
Dans le monde diurne, le système rétinien le plus évolué pour discriminer les détails fins
d’un objet est constitué par une fovéa ; elle est présente chez les lézards, les primates.
On en trouve parfois deux chez certains oiseaux. L’aigle dispose d’un système optique
d’agrandissement et de deux fovéas : une fovéa monoculaire, centrale, qui observe et
une seconde fovéa binoculaire, temporale, qui permet un guidage très précis de
l’approche et de la capture de la proie entre les pattes.
Area centralis
Une autre particularité de la rétine des vertébrés est l’absence de macula, à l’exception
des primates non humains. Il existe cependant une zone appelée area centralis [Peichl,
1992] où la proportion de cônes est plus importante qu’ailleurs mais l’area centralis ne
présente pas de repères topographiques permettant de l’identifier par un examen
ophtalmoscopique.
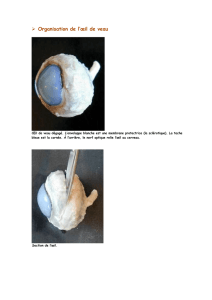
La figure VIII-5 montre la régionalisation fonctionnelle de la neurorétine chez le chien.
Dans la partie ventrale, l’épithélium pigmentaire est pigmenté alors que dans la partie
dorsale, l’épithélium pigmentaire est dépigmenté, révélant la structure choroïdienne
appelé le Tapetum lucidum.
L’area centralis se situe dans la partie dorsolatérale à quelques millimètres au dessus de
la papille. Son diamètre est d’environ 0.2 mm chez le chien. Il n’y a pas de fovéa à
proprement parler car des bâtonnets y sont toujours présents. En cette zone centrale, les
cônes sont plus petits et plus minces qu’en rétine périphérique.
La densité des cônes y est d’environ 27 000/mm² ; elle décroît de façon importante en
périphérie pour atteindre une valeur d’environ 4 000/mm².
La densité des bâtonnets est sensiblement constante de l’ordre de 250 000/mm² sauf
autour de l’area centralis, où il existe une zone de densité maximale des bâtonnets au
environ de 450 000/mm².
Animaux nocturnes
Au contraire, la rétine des animaux nocturnes (poissons abyssaux, oiseaux, la plupart
des rongeurs de laboratoire) comporte essentiellement des bâtonnets qui favorisent leur
sensibilité à la lumière et aux mouvements, en ambiance scotopique [Szel, Rohlich,
1992], [Peichl, 2005], [Mustafi et al., 2009].
Une structure choroïdienne particulière : le Tapetum
Lucidum
Le fonctionnement de la rétine de tous les vertébrés est à peu près identique mais, d’un
point de vue ophtalmoscopique, les aspects des fonds d’yeux et de leur vascularisation
présentent de grandes variations. Elles sont en particulier liées à la présence d’une
structure choroïdienne spécifique : le Tapetum lucidum.
Structure choroïdienne, le Tapetum Lucidum est absent chez les rongeurs, le porc et les
primates et revêt de nombreuses variations chez les autres espèces.
Différents aspects du Tapetum Lucidum
Il représente plus des deux-tiers de la surface rétinienne chez le chat. Chez les
carnivores, il est de type cellulaire et sa composition est différente selon l’espèce. Chez le
chien, il est très riche en zinc-cystéïne [Lesiuk, Braekevelt, 1983], [Chijiiwa et al., 1990]
alors que chez le chat, il est essentiellement composé de riboflavines [Braekevelt, 1990].
Chez les ongulés, il est de type fibreux [Braekevelt, 1986].
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
1
/
47
100%