Chapitre 2.1.13.

CHAPITRE 2.1.13.
PESTE PORCINE CLASSIQUE
(hog cholera)
RÉSUMÉ
La peste porcine classique (PPC), aussi connue sous la dénomination de hog cholera, est une
maladie virale contagieuse du porc. Le virus responsable est un membre du genre Pestivirus de la
famille des Flaviviridae, et se révèle très proche des virus de la maladie des muqueuses et de la
maladie de la frontière (border disease). Il n’existe qu’un sérotype de virus PPC.
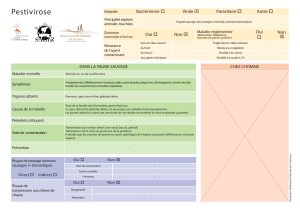
La maladie peut évoluer sous forme aiguë, chronique, latente, ou inapparente selon divers facteurs
tenant au virus ou à l’hôte, parmi eux l’âge de l’animal, la virulence de la souche virale et la période
d’infection (pré- ou post-natale) jouent un rôle prépondérant. Les porcs adultes sont moins
sévèrement atteints que les jeunes et ont de meilleures chances de survie. Chez les truies
gestantes, le virus peut traverser la barrière placentaire et atteindre les fœtus. L’infection in utero
par des souches virales de basse ou faible virulence peut aboutir à ce que l’on a appelé le
syndrome de la « la truie porteuse », entraînant une mortalité pré-natale ou post-natale précoce, la
naissance de porcelets malades ou d’une portée en apparente bonne santé mais infectée. Une
épizootie de PPC a des répercussions lourdes sur le commerce des porcs et de leurs produits.
Le tableau clinique très polymorphe de la PPC interdit le plus souvent la réalisation d’un diagnostic
purement clinique ou lésionnel. Les méthodes de laboratoire sont alors essentielles pour un
diagnostic précis. La mise en évidence du virus dans le sang et des anticorps dans le sérum,
constituent les méthodes de choix du diagnostic de la PPC chez le porc vivant, tandis que la
recherche du virus ou de l’antigène dans des prélèvements d’organes, est de loin préférable quand
le porc est mort.
Identification de l’agent pathogène : L’immunofluorescence (IF) appliquée sur des coupes au
cryostat d’organes provenant de porcs atteints, est utilisée pour la détection de l’antigène PPC.
Une série d’anticorps monoclonaux (AcMs) est utilisée pour déterminer si la fluorescence est due à
des antigènes pestivirus PPC ou non PPC. L’isolement du virus PPC peut être tenté sur la lignée
cellulaire de rein de porc (PK-15) ou sur une autre lignée sensible. La production virale en culture
de cellules est recherchée par coloration en immunofluorescence ou immunoperoxydase ; les
isolats positifs sont ensuite caractérisés grâce aux AcMs et par un séquençage génétique partiel.
Les protocoles des réactions d’amplification en chaîne par polymérase (PCR) pour l’identification
de l’acide nucléique du virus PPC sont utilisés dans plusieurs laboratoires. L’isolement et
l’identification des souches virales pathogènes suspectes doivent être conduits dans des
laboratoires répondant aux normes de sécurité.
Épreuves sérologiques : La recherche des anticorps spécifiques du virus est particulièrement
utile dans les élevages suspectés d’être infectés depuis au moins 30 jours, par le virus de la PPC.
Les méthodes sérologiques sont aussi valables pour les enquêtes de contrôle et de détermination
de prévalence ; elles sont essentielles si un pays souhaite être reconnu internationalement comme
indemne de la maladie en l’absence de vaccination.
Dans la mesure où des réactions croisées avec des anticorps Pestivirus de ruminants sont parfois
rencontrées chez les porcs d’élevage, les épreuves de dépistage doivent être complétées par des
épreuves de confirmation spécifiques du virus PPC. L’ELISA est relativement spécifique du virus
PPC, mais la méthode définitive de différentiation est l’épreuve de neutralisation comparative qui
compare le titre en anticorps vis-à-vis des différentes espèces de Pestivirus.
274 Manuel terrestre de l’OIE 2005

Chapitre 2.1.13. — Peste porcine classique (hog cholera)
Spécifications applicables aux vaccins et aux produits biologiques à usage diagnostique :
Les vaccins contre la PPC utilisent des virus vivants atténués par passages sur cultures de cellules
ou sur des espèces réceptives qui ne sont pas de la famille des suidés. La production de ces
vaccins à virus vivants modifiés repose sur la validation du système lot-semence tenant compte de
l’identité virale, de la stérilité, de la pureté, de l’innocuité, de la non transmissibilité, de la stabilité et
de l’immunogénicité. Si le virus PPC est utilisé pour la production de vaccin ou lors d’épreuves
virulentes, les locaux doivent répondre aux normes de confinement de l’OIE correspondant aux
agents pathogènes du groupe 4.
Des vaccins inactivés de type conventionnel, impliquant le virus entier, ne sont pas disponibles.
Ces dernières années des « vaccins avec marqueur sérologique » ont été développés, qui, à la
différence des vaccins à virus vivants atténués, induisent des anticorps qui peuvent être distingués
de ceux induits par le virus naturel en utilisant une épreuve de diagnostic appropriée. Les « vaccins
avec marqueur sérologique » proposés à l’heure actuelle, utilisent la glycoprotéine majeure de
l’enveloppe (E2-sous-unité) du virus PPC et sont produits sur cellules d’insecte grâce à la
méthodologie de recombinaison de l’ADN.
A. INTRODUCTION
Les virus responsables de la peste porcine classique (PPC), de la diarrhée virale bovine (BVD, Bovine Viral
Diarrhoea ou maladie des muqueuses) et de la maladie de la frontière (BD, Border Disease) sont des membre de
la famille des Flaviviridae, appartenant au genre Pestivirus, et sont très proches au plan antigénique et structurel.
Les signes cliniques et les lésions découvertes à l’autopsie de porcs morts de PPC sont très variables en raison
de facteurs liés, à la fois au virus et à l’hôte. Bien plus, les infections congénitales de la truie par des pestivirus de
ruminants peuvent conduire à une maladie cliniquement indistinguable de la PPC (22, 24, 25).
L’atteinte de tous les groupes d’âge, accompagnée de fièvre, d’entassement, d’inappétence, d’abattement, de
faiblesse, de conjonctivite, de constipation suivie par une diarrhée et d’une démarche chancelante, sont les
signes essentiels. Plusieurs jours après l’apparition des signes cliniques, une coloration pourpre apparaît sur les
oreilles, l’abdomen et à l’intérieur des cuisses. La mort survient en 1 à 2 semaines dans la forme aiguë. Une mort
brutale, en l’absence de signes cliniques, n’est pas symptomatique de la PPC.
Dans certaines circonstances en rapport avec l’âge de l’animal et son état ainsi qu’avec la souche de virus en
cause, des atteintes subaiguë ou chronique peuvent se développer et se prolonger sur 2 à 4 semaines ou même
des mois. La forme chronique conduit à un retard de croissance avec anorexie, fièvre intermittente et diarrhée.
Les infections congénitales persistantes peuvent restées méconnues pendant des mois et peuvent être limitées à
seulement quelques porcelets de l’élevage. Les signes cliniques sont équivoques : dépérissement apyrétique.
Les infections chroniques persistantes conduisent toujours à la mort de l’animal. Les taux de mortalité dans un
élevage, peuvent être légèrement supérieurs à ceux attendus. La PPC affecte le système immunitaire et la
principale caractéristique en est une leucopénie généralisée, qui peut souvent être détectée avant l’apparition de
la fièvre. L’immunosuppression peut entraîner l’apparition d’infections secondaires.
Dans les formes aiguës, les lésions macroscopiques sont souvent discrètes ou absentes. Dans les cas typiques,
les nœuds lymphatiques sont hypertrophiés et marbrés de rouge, des hémorragies apparaissent sur l’épicarde et
dans les reins, la vessie, la peau et le conjonctif sous-cutané. Dans les cas chroniques, des ulcères nécrotiques
dits « en boutons de culotte » peuvent être observés dans la muqueuse du tractus gastro-intestinal, l’épiglotte et
le larynx, en plus des lésions précédentes.
Les lésions microscopiques histologiques ne sont pas pathognomoniques. Il peut s’agir de dégénérescence
parenchymateuse du tissu lymphatique, de proliférations cellulaires du tissu interstitiel vasculaire, et d’une
méningo-encéphalomyélite non suppurée, avec ou sans manchons péri-vasculaires.
B. TECHNIQUES DE DIAGNOSTIC
Le polymorphisme et l’intensité variable des signes cliniques et lésionnels ne permettent pas de conduire un
diagnostic précis. D’autres maladies, telles que la Peste porcine africaine, les troubles du sevrage, le syndrome
de dépérissement et le syndrome dermatite et néphropathie du porc, ainsi que des infections septicémiques telles
que les salmonellose, pasteurellose, actinobacillose et les infections à Hemophilus suis, peuvent être confondues
avec la PPC aiguë. En fait, ces bactéries sont souvent responsables d’infections secondaires et, l’isolement de
ces germes peut cacher la cause réelle de la maladie, le virus de la PPC.
Manuel terrestre de l’OIE 2005 275

Chapitre 2.1.13. — Peste porcine classique (hog cholera)
C’est pourquoi, une tentative de diagnostic clinique ou lésionnel doit toujours être confirmée par des recherches
de laboratoire. Cela est d’autant plus nécessaire à la vue des lourdes conséquences que l’existence d’un foyer de
PPC peut causer pour le commerce international des porcs et de leurs produits.
Les méthodes de laboratoire pour le diagnostic de la PPC visent la mise en évidence du virus, de son acide
nucléique ou de ses antigènes ainsi que des anticorps qui lui sont spécifiques. Pour une interprétation correcte
des résultats des épreuves, le vétérinaire doit accorder une attention particulière à l’apparition simultanée et
groupée d’au moins 2 signes parmi les plus fréquents de la maladie signalés ci-dessus. La réalisation de
prélèvements au hasard ne convient pas au diagnostic. Etant donné que la fièvre est un des premiers signes de
PPC et s’accompagne d’une virémie (6), la recherche du virus dans le sang récolté sur acide éthylène-diamine-
tétra-acétique (EDTA) ou dans des tissus, prélevés sur un animal fiévreux, est la méthode de choix pour détecter,
précocement, les élevages infectés. De plus, les prélèvements sanguins destinés à la recherche du virus peuvent
être réalisés sur un nombre plus important de porcs.
La PPC fait l’objet d’un contrôle officiel et le virus présente un haut risque de diffusion à partir du laboratoire : une
analyse de risque doit être effectuée pour déterminer le niveau de biosécurité nécessaire au diagnostic et à la
caractérisation du virus. Les locaux doivent répondre aux normes de Confinement correspondant au Groupe
déterminé par l’évaluation du risque comme cela est souligné dans l’appendice I.1.6.1. du Chapitre I.1.6., « La
biosécurité au laboratoire de microbiologie vétérinaire », de ce Manuel terrestre. Les pays ne pouvant avoir accès
à un tel laboratoire national ou régional, devront adresser les prélèvements à un Laboratoire de référence de
l’OIE.
Les anticorps apparaissent durant la troisième semaine d’évolution de la maladie et persistent toute la vie chez
les sujets survivants. Les prélèvements destinés à la recherche des anticorps sont effectués dans des tubes
ordinaires non héparinés, chez les porcs convalescents et chez les porcs de l’élevage qui ont été en contact avec
le virus, au moins 30 jours après l’apparition suspectée du virus.
1. Identification de l’agent pathogène
a) Méthodes immunologiques
• Épreuve d’immunofluorescence
L’immunofluorescence (IF) est rapide et peut permettre de détecter l’antigène du virus PPC sur des coupes
au cryostat d’amygdale, de rate, de rein, de nœud lymphatique ou de portion distale de l’iléon. Les tissus
doivent être prélevés sur plusieurs animaux (3) et transportés sans conservateur, sous froid mais non
congelés. Les coupes au cryostat sont colorées directement avec une immunoglobuline anti-PPC conjugué
à l’isothiocyanate de fluorescéine (FITC) ou indirectement à l’aide d’un conjugué FITC secondaire puis
examinées au microscope à fluorescence. Au début de l’infection, le tissu amygdalien est le plus propice car
il est le premier à être infecté par le virus quelle que soit la voie de pénétration (18). Dans les cas subaiguës
ou chroniques, l’iléon est souvent positif et, parfois, constitue le seul tissu révélant une fluorescence. Un
résultat négatif en IF ne peut permettre d’éliminer l’infection PPC. Lorsque la suspicion de PPC persiste, il
doit être réalisé d’autres prélèvements ou des essais d’isolement du virus en cultures de cellules (par
exemple sur rein de porc [PK-15] ou sur d’autres lignées d’origine porcine aussi sensibles et reconnues
exemptes de contamination par Pestivirus).
• Protocole
Introduire des coupes témoins positif et négatif dans chaque série de prélèvements d’organe à examiner.
i) Découper un fragment d’amygdale, de rate, de rein et d’iléon d’environ 1 x 1 x 0,5 cm, et le placer soit
avec un composé de cryo-montage, soit avec de l’eau distillée, sur le support de tissu du cryostat.
ii) congeler le fragment d’organe sur le support du cryostat.
iii) Effectuer des coupes ne dépassant pas 4 µm d’épaisseur et les étaler sur des lamelles couvre-objet
dont un coin est coupé. Toutes les coupes sont montées avec le coin coupé de la lamelle dans la
même direction (par exemple angle supérieur droit).
iv) Après séchage, fixer les coupes ainsi montées, pendant 10 min à la température ambiante, dans de
l’acétone (qualité analytique), ou à l’air pendant 20 min à 37°C.
v) Immerger rapidement les coupes dans une solution physiologique tamponnée au phosphate (PBS),
éliminer le fluide en excès à l’aide d’un papier filtre, et les placer (angle supérieur coupé à droite) sur
un support dans une chambre humide dont le fond renferme une petite quantité d’eau.
276 Manuel terrestre de l’OIE 2005

Chapitre 2.1.13. — Peste porcine classique (hog cholera)
vi) Répartir l’immunoglobuline anti-PPC à la dilution recommandée, sur la totalité de la surface de la
coupe et incuber pendant 30 min à 37°C dans la chambre humide close. Si le conjugué FITC
secondaire est nécessaire, laver la coupe 5 fois, pendant 2 min chacune, dans du PBS à la
température ambiante, puis ajouter le conjugué FITC à la dilution requise et incuber comme
précédemment.
vii) Laver les coupes 5 fois, pendant 2 min chacune, dans du PBS, à la température ambiante.
viii) Eliminer le PBS restant par contact de la lamelle avec un papier filtre fin, et monter la lamelle (avec la
coupe entre la lamelle et la lame) avec un tampon de montage, sur une lame pour examen
microscopique.
ix) Eliminer l’excès de fluide de montage avec du papier filtre et examiner les coupes pour rechercher les
foyers fluorescents, en utilisant un microscope à lumière ultra-violette. Une coupe PPC-positive montre
des cellules émettant une fluorescence verte brillante. Dans les amygdales, la fluorescence de la
bordure épithéliale des cryptes est particulièrement marquée. Dans le rein, la fluorescence est plus
intense dans les tubules proximaux et distales du cortex rénal et les tubes collecteurs de la médulla.
Dans l’iléon, la fluorescence est plus accusée dans les cellules épithéliales des glandes de Lieberkün,
tandis que dans la rate, la réactivité est plus diffuse avec des concentrations de cellules lymphoïdes
dans les gaines lymphoïdes péri-artérielles (PALS).
L’IF utilise une immunoglobuline anti-PPC préparée à partir d’un anticorps polyclonal du virus PPC
incapable de distinguer les antigènes des différents pestivirus. Les conjugués utilisés pour l’IF sur les
coupes au cryostat ou les cultures de cellules inoculées, doivent être élaborés à l’aide de gamma-globulines
obtenues chez des porcs exempts d’agents pathogènes spécifiques (EAPS). La dilution de conjugué utilisée
(au moins 1/30) doit ménager un maximum de brillance sur un minimum de coloration de fond.
Les souches vaccinales de virus vivant modifié (VVM) se multiplient surtout dans les nœuds lymphatiques
régionaux et dans l’épithélium des cryptes amygdaliennes. Les porcs vaccinés avec des souches VVM
peuvent produire une IF positive durant les 2 semaines suivant la vaccination (15, 19). L’inoculation du lapin
est utilisée pour différencier les souches de PPC lapinisées de celles du terrain. A l’opposé des souches de
terrain, les souches lapinisées, inoculées par voie intraveineuse, entraînent une réaction fébrile et induisent
une réponse immunitaire chez le lapin.
Les porcs infectés par les pestivirus des ruminants peuvent donner, en IF, des réactions faussement
positives. Les infections congénitales dues aux pestivirus des ruminants peuvent entraîner des signes
cliniques et des lésions identiques à ceux et à celles de la PPC chronique (22, 24, 25). Les infections par le
virus PPC ou les pestivirus des ruminants peuvent être différenciés en soumettant les sérums de la truie et
de la portée, ou des autres sujets en contact avec un porcelet IF positif, à une épreuve de séroneutralisation
vis-à-vis de chacun des virus. Une autre méthode de différentiation de ces virus consiste en l’inoculation de
porcelets séronégatifs avec une suspension du matériel suspect, suivie, 5 semaines plus tard, d’épreuves
de séroneutralisation pratiquées sur leur sérums pour la recherche des anticorps respectifs. Cependant, les
épreuves de séroneutralisation peuvent prendre plusieurs jours et les méthodes d’inoculation à l’animal
réclament plusieurs semaines.
• Épreuve d’immunoperoxydase pour la différentiation des pestivirus par les anticorps monoclonaux
Le recours à une série de 3 anticorps monoclonaux (AcMs), soit conjugués à la peroxydase de raifort
(HRPO) ou à l’isothiocyanate de fluorescéine (FITC), soit utilisés en association avec un conjugué anti-
souris et détectant de façon spécifique respectivement toutes les souches PPC de terrain, les souches
vaccinales PPC et les pestivirus des ruminants, permettrait une différentiation précise entre les souches de
terrain et vaccinales PPC, d’une part, et entre virus PPC et les autres pestivirus, d’autre part (10, 26, 28).
Une condition essentielle est que l’AcM anti-virus PPC reconnaisse toutes les souches terrain et que l’AcM
anti-vaccin reconnaisse toutes les souches vaccinales utilisées dans le pays. Aucun AcMs ne réagit
sélectivement avec tous les pestivirus de ruminants (10). Le recours à un AcM différenciant la souche
vaccinale PPC n’est pas nécessaire dans les zones où l’on ne vaccine pas. Une immunoglobuline
polyclonale anti-PPC conjuguée à la peroxydase (HRPO) sert de témoin positif. Des précautions doivent
être prises lorsque l’on utilise un seul AcM en confirmation unique de la nature PPC d’un isolat.
• Protocole
i) Réaliser au cryostat au moins 8 coupes (4 µm) d’amygdale positive en IF ou d’un autre organe positif
si l’on ne dispose pas d’amygdale.
ii) Fixer les coupes sur des lamelles couvre objet dans l’acétone (qualité analytique) pendant 10 min et
laisser sécher à l’air.
Manuel terrestre de l’OIE 2005 277

Chapitre 2.1.13. — Peste porcine classique (hog cholera)
iii) Préparer des dilutions appropriées des différents conjugués AcMs-peroxydase en PBS + 0,01 % de
Tween 80 + 5 % de sérum de cheval, pH 7,6 (un FITC-AcMs peut aussi être utilisé, aussi bien qu’un
AcM non conjugué, pourvu qu’un second conjugué soit utilisé).
iv) Après rinçage en PBS, recouvrir 2 coupes avec la dilution des conjugués monoclonaux respectifs et
2 coupes avec la dilution du conjugué polyclonal (témoins).
v) Incuber à 37°C pendant 1 h en chambre humide.
vi) Laver les coupes pendant 10 s, 6 fois, en PBS.
vii) Colorer les coupes avec la solution de substrat chromogène∗ fraîchement préparée, pendant 5 à
15 min à température ambiante.
viii) Rincer les coupes dans une solution en eau distillée d’acétate de sodium 0,05 M, pH 5,0, et les monter
sur des lames pour examen microscopique.
ix) Examiner les coupes au microscope. Une coloration rouge foncé du cytoplasme des cellules
épithéliales bordant les cryptes amygdaliennes indique la mise en évidence d’un virus par le conjugué
correspondant et est considéré positive.
x) Interprétation de l’épreuve :
Anticorps
polyclonal Anticorps monoclonal spécifique de Interprétation
Souche PPC Souche vaccinale
PPC Souche BVD/BD
+ + – –
Souche terrain PPC
+ + + – Souche vaccinale PPC
+ – – + Souche BVD/BD
+ – – –
Autre pestivirus non-PPC*
* L’existence de nouvelles souches de PPC doit toujours être pris en compte et tout isolat de cas où la PPC est
encore suspectée doit être envoyé à un Laboratoire de référence de l’OIE.
• Méthode immuno-enzymatique de capture
Pour un diagnostic rapide de la PPC chez des porcs vivants, des épreuves immuno-enzymatique (ELISA)
de capture ont été mises au point pour détecter les élevages suspects d’avoir été récemment infectés. Les
ELISAs sont du type sandwich double-anticorps, avec des anticorps monoclonaux et/ou polyclonaux dirigés
contre diverses protéines virales et utilisables sur du sérum, la fraction leucocytaire sanguine, ou du sang
total prélevé sur anticoagulant (des homogénats de tissu centrifugés peuvent être ajoutés ici dans la mesure
où ils constituent un matériel utilisable en ELISA) (7). La technique est relativement simple à effectuer, ne
réclame pas d’équipement nécessaire à la culture de tissu, est automatisable et peut donner des résultats
en une demi-journée. L’inconvénient d’être moins sensible que l’isolement viral, spécialement chez le porc
adulte et lors de formes modérées ou sub-cliniques, est compensé par la possibilité de tester tous les porcs
fiévreux suspects d’un élevage. Néanmoins, la faible spécificité de ces épreuves doit aussi être considérée.
b) Isolement du virus
L’isolement du virus en cultures de cellules est une méthode plus sensible mais plus lente pour le diagnostic
de la PPC que l’immunofluorescence sur coupes en congélation. L’isolement est, au mieux, réalisé en
cellules à division rapide PK-15 ensemencées sur des lamelles simultanément avec une suspension
d’amygdale à 2 % dans le milieu de culture. D’autres lignées de cellules de porc peuvent être utilisées, mais
elles doivent avoir démontré qu’elles sont au moins aussi sensibles que les cellules PK-15 pour l’isolement
du virus PPC. Les cultures sont examinées par IF pour recherche des foyers fluorescents, après 24 à 72 h.
∗ Solution de substrat chromogène
A. Solution stock de chromogène : 0.4 % de 3-amino-9-ethyl carbazole; N,N-dimethyl-formamide (1 ml).
Prendre garde aux composés toxiques.
B. 0,05 M d’acétate de sodium, pH 5,0 ; 19 ml (filtré stérilement à travers une membrane).
C. Solution stock de substrat (30 % de peroxyde d’hydrogène).
Conserver les solutions stock A et C dans l’obscurité et à 4°C et la solution B à température ambiante. La solution stock A peut
être conservée à 4°C Durant au moins 6 mois et la solution C durant au moins 1 an. Juste avant d’utiliser les solutions, diluer
1 ml de la solution A dans 19 ml de la solution B. Ajouter alors 10 µl de la solution stock C. Mélanger bien et colorer les coupes.
278 Manuel terrestre de l’OIE 2005
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%