MÉMOIRE L`effet préventif de quelques huiles essentielles contre la

République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
Université Aboubekr Belkaïd de Tlemcen
Faculté des Sciences de la Nature et de la Vie et des Sciences de la Terre et de l’Univers
Département de Biologie
Laboratoire de Microbiologie Appliquée à l’Agroalimentaire, au Biomédical et à l’Environnement (LAMAABE)
MÉMOIRE
pour l’obtention du diplôme de Magister en Biologie
option : Maîtrise de la Qualité Microbiologique et du Développement Microbien
Thème
L’effet préventif de quelques huiles essentielles contre la
croissance et la formation de biofilms de certains pathogènes de
la cavité buccale
par
Mr. Mohamed Amine Abdoune
soutenu le 23/05/2013
devant le jury composé de
Pr. Djamel Eddine Abdelouahid Président Professeur UABT
Dr. Hafida Hassaïne Examinatrice Maître de conférences classe A UABT
Dr. Hamadi Abderrahmane Lazouni Examinateur Maître de conférences classe A UABT
Dr. Mourad Bendahou Promoteur Maître de conférences classe A UABT
Mme. Fadila Malek Invité Maître assistant chargée de cours UABT
Année universitaire 2012‒2013

République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
Université Aboubekr Belkaïd de Tlemcen
Faculté des Sciences de la Nature et de la Vie et des Sciences de la Terre et de l’Univers
Département de Biologie
Laboratoire de Microbiologie Appliquée à l’Agroalimentaire, au Biomédical et à l’Environnement (LAMAABE)
MÉMOIRE
pour l’obtention du diplôme de Magister en Biologie
option : Maîtrise de la Qualité Microbiologique et du Développement Microbien
Thème
L’effet préventif de quelques huiles essentielles contre la
croissance et la formation de biofilms de certains pathogènes de
la cavité buccale
par
Mr. Mohamed Amine Abdoune
soutenu le 23/05/2013
devant le jury composé de
Pr. Djamel Eddine Abdelouahid Président Professeur UABT
Dr. Hafida Hassaïne Examinatrice Maître de conférences classe A UABT
Dr. Hamadi Abderrahmane Lazouni Examinateur Maître de conférences classe A UABT
Dr. Mourad Bendahou Promoteur Maître de conférences classe A UABT
Mme. Fadila Malek Invité Maître assistant chargée de cours UABT
Année universitaire 2012‒2013

Dédicace
Je dédie ce modeste travail
à mes précieux parents
et à mon cher frère
Mohamed Amine

Remerciements
Ce travail à été réalisé au Laboratoire de Microbiologie Appliquée à l’Agroalimentaire, au
Biomédical et à l’Environnement (LAMAABE) sous la direction de monsieur M. Bendahou, maître
de conférences classe A au Département de Biologie, Faculté des Sciences de la Nature et de la Vie et
des Sciences de la Terre et de l’Univers (SNV-STU), Université de Tlemcen. Je tiens à lui formuler
mes profondes reconnaissances et gratitudes pour l’honneur qu’il a fait d’avoir accepté de diriger ce
mémoire. Je le remercie pour l’aide et les conseils précieux qu’il n’a cessé de prodiguer tout le long
du travail.
Je tiens à remercier monsieur D. E. Abdelouahid, professeur au Département de Biologie,
Faculté des SNV-STU, Université de Tlemcen d’avoir accepté de présider le jury.
Je formule mes sincères gratitudes à madame H. Hassaïne, maître de conférences classe A au
Département de Biologie, Faculté des SNV-STU, Université de Tlemcen d’avoir accepté de juger ce
travail.
Je remercie également monsieur H. A. Lazouni, maître de conférences classe A au
Département d’Écologie, Faculté des SNV-STU, Université de Tlemcen d’avoir accepté d’être parmi
les jurés afin d’examiner ce travail.
J’exprime infiniment mes fortes reconnaissances à madame F. Malek, maître assistant chargée
de cours au Département de Biologie, Faculté des SNV-STU, Université de Tlemcen pour son aide,
ses précieux conseils et pour la chance qu’elle m’a offerte en assistant les manipulations sur les
biofilms agroalimentaires.
Sans oublier aussi monsieur le directeur du laboratoire LAMAABE, le professeur B. Moussa-
Boudjemâa pour tous ces conseils et son esprit de communication avec tous les gens au sein du
laboratoire.
Je remercie aussi tous mes collègues au laboratoire étudiants et enseignants, et en particulier,
F. Benbelaïd et A. Khadir, pour leur aide continu tout au long du travail, je les félicite de leur esprit
d’équipe et je leur souhaite une bonne continuation.
Pour n’oublier aucune personne, je formule mes sincères remerciements à tous les gens qui
ont participé de près ou de loin à la réalisation de ce modeste travail.

Résumé
L’effet préventif de quelques huiles essentielles contre la croissance et la formation de biofilms de
certains pathogènes de la cavité buccale.
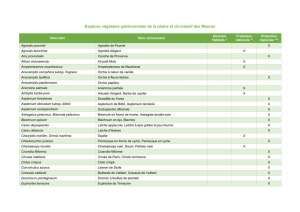
Les huiles essentielles de vingt espèces végétales récoltées dans la région de Tlemcen ont été
testées sur la croissance et la formation de biofilms de cinq souches microbiennes connues par leur
pathogénicité dans la cavité buccale. La plupart des huiles essentielles étaient actives surtout sur les
bactéries à Gram positif et sur Candida albicans. Les huiles essentielles les plus actives étaient celles
de Thymbra capitata (CMI = 0,31 mg/ml pour C. albicans) et d’Origanum vulgare subsp.
glandulosum (CMI = 0,63 mg/ml pour S. aureus et E. faecalis). Les CMI de la formation de biofilms
étaient généralement très faibles par rapport à celles de la croissance sauf pour C. albicans qui
étaient semblables.
Mots-clés : huile essentielle ; croissance ; formation de biofilm ; pathogènes ; cavité buccale.
Abstract
Preventive effect of some essential oils against growth and biofilm formation of some oral
pathogens.
Essential oils of twenty plant species harvested in the Tlemcen region were tested on growth
and biofilm formation of five microbial strains known for their pathogenicity in the oral cavity.
Most essential oils were active mainly on Gram-positive bacteria and Candida albicans. The most
active essential oils were those of Thymbra capitata (MIC = 0.31 mg/ml for C. albicans) and
Origanum vulgare subsp. glandulosum (MIC = 0.63 mg/ml for S. aureus and E. faecalis). The MICs
of biofilm formation were generally quite low compared to those of growth except for C. albicans
which were similar.
Keywords: essential oil; growth; biofilm formation; pathogens; oral cavity.
صخلم
!"#$%&'()*+,-./.012
3.4
5"6789-:95+*!;!.:<2&!"!=>7?'@AB.9<C"9!D-2>7
E.+!),'(%&#$FG"C,H9.9IJKL".9I*. !M3.4NL;O9
CP'".2&IQR"S.,T/S.U.05JVL.CNL
WXY7893Q3Z[0,319\]9S.*^/S._(,"#R[3Q3Z0,639\]989(-:
NL3^=`,".JK.LYV,. 1.4<S$a:9>7QD-,2b9F*!Pa
3*9NLU.0/S.S.,/:VB2:2
*cc#7N=d3#C%&!c)*+9;.<c.0 !
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
1
/
109
100%