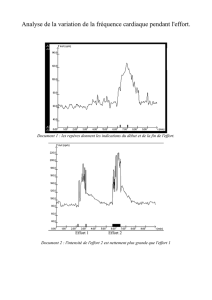
Physiologie animale

Sherwood
I
Klandorf
I
Yancey
Sherwood
I
Klandorf
I
Yancey
Physiologie
animale
Physiologie animale
L’ouvrage traite, de façon exhaustive, des différentes
fonctions qui sous-tendent la vie des animaux. Chacune
d’elles est appréhendée à l’échelle des organismes,
dans ses manifestations visibles extérieurement (com-
portementales notamment), mais aussi à l’échelle des
appareils et organes qui en sont les sites d’expression
et à celle des cellules — avec une incursion pointue
dans les signalisations intracellulaires mises en jeu, les
intermédiaires moléculaires sollicités, les gènes qui en
détiennent l’information et les contrôles qui s’exercent
dans le cadre du maintien de l’homéostasie.
Des contenus accessibles, actuels et exacts
L’ouvrage est conçu selon un format logique et est
enrichi de multiples exemples. De l’information perti-
nente basée sur des découvertes récentes, ainsi que de
nouvelles méthodologies de recherche ont été incluses
dans chacun des chapitres. Des idées controversées et
des hypothèses sont également présentées pour illustrer
le fait que la physiologie est une discipline dynamique
et évolutive.
De la globalité à la spécifi cité
La physiologie est replacée dans le contexte de l’évo-
lution animale, avec le souci permanent de mettre
l’accent sur le fait que chaque concept est un élément
incontournable qui s’intègre dans le sujet considéré dans
sa globalité. Des efforts particuliers ont été faits pour
assurer une lecture fl uide avec des transitions aisées,
un raisonnement logique et l’intégration d’idées clés et
fondamentales à travers l’intégralité de l’ouvrage.
Traduction de la seconde édition américaine
par Jean-Pierre Cornec
Formateur dans la préparation aux concours du CAPES
et de l’Agrégation des Sciences de la vie, de la Terre et
de l’Univers. Ancien enseignant, chercheur et maître de
conférences.
Physiologie animale
Sherwood
I
Klandorf
I
Yancey
a Physiologie comparée et évolutive étudiée sur
l’ensemble du monde animal
a Illustrations abondantes et diversifi ées, en couleurs
a Bibliographie à la fi n de chaque chapitre avec
commentaires
a Couvre une bonne partie de l’enseignement de la
physiologie animale du premier cycle
Zoologie
Traduction de Jean-Pierre Cornec
Miller
I
Harley
Chez le même éditeur
http://noto.deboecksuperieur.com : la version numérique de votre ouvrage
• 24h/24, 7 jours/7
• O ine ou online, enregistrement synchronisé
• Sur PC et tablette
• Personnalisation et partage
9 782807 302860
ISBN : 978-2-8073-0286-0
Conception graphique : Primo&Primo®
illu : © ilbusca / decade3d
SHERWOODANI
Traduction de Jean-Pierre Cornec
9782807302860_SHERWOODANI_CV.indd 1 02/11/2016 12:27


Physiologie
animale
68651_PC_CS4.indb 1 28/10/16 11:34

Chez le même éditeur
BERG, HASSENZAHL & RAVEN, Environnement.
BERTHET, Dictionnaire de biologie.
CAIN, DAMMAN, LUE & YOON, Découvrir la biologie.
FORÊT, Dico de Bio.
GAUTHIER-CLERC, Sciences de la conservation.
ANCTIL, BAGUET, CHARMANTIER, GILLES, GILLES JR, PÉQUEUX, PLUMIER, SÉBERT, Physiologie animale.
HILBER, Physiologie animale.
HARLEY, MILLER, Zoologie.
RAVEN, JOHNSON, MASON, LOSOS, & SINGER, Biologie.
TANZARELLA, Perception et communication chez les animaux.
TAUTZ, L’étonnante abeille.
68651_PC_CS4.indb 2 28/10/16 11:34

Sherwood,
Klandorf, Yancey
Physiologie
animale
Traduction de la seconde édition américaine
par Jean-Pierre Cornec
68651_TDM.indd 3 02/11/16 13:56
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
1
/
60
100%