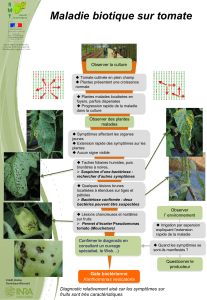
Verticillium

221
UNIVERSITE MOHAMMED V- Agdal
FACULTE DES SCIENCES
RABAT
THESE DE DOCTORAT D’ETAT
Présentée par
Amina REGRAGUI
Discipline : Biologie
Spécialité : Phytopathologie
Contribution à l’étude de l’influence de la salinité sur
le couple tomate –Verticillium : Conséquences
physiologiques et impact sur la bioprotection des
tomates contre la verticilliose
Soutenue le…22 décembre 2005…..devant le jury :
LAHLOU H.
Professeur à la Faculté des Sciences de Rabat Présidente
BENKHEMMAR O.
Professeur à la Faculté des Sciences de Rabat Examinateur
FASSI FIHRI O.
Professeur à l’I.A.V. Hassan II, Rabat Examinateur
TIJANE M.
Professeur à la Faculté des Sciences de Rabat Examinateur
ZAID H.
Professeur à la Faculté des Sciences de Rabat Examinateur

231
JE DEDIE CE TRAVAIL…
A mes petits enfants
Zineb
Youssef
Jad
et……

229
REMERCIEMENTS
Ce travail a été réalisé au sein de l’équipe de phytopathologie
du laboratoire de Botanique de la Faculté des Sciences de Rabat.
Qu’il me soit permis ici de remercier vivement le professeur H.
LAHLOU pour la confiance qu’elle m’ a toujours témoignée ainsi que
la formation de chercheur qu’elle m’a donnée avec bienveillance et
humanité. Ses qualités scientifiques, ses conseils et ses
encouragements incessants m’ont beaucoup soutenue pendant la
l’élaboration de ce travail. Je lui exprime ma profonde
reconnaissance et lui porte mon grand respect.
Je remercie très vivement le professeur H. ZAID pour sa
disponibilité, ses conseils fructueux et son aide scientifique et
matérielle. Il a bien voulu accepter de participer à ce jury de thèse
en qualité de rapporteur. Qu’il trouve ici l’’expression de ma
profonde gratitude.
Madame O. FASSI FIHRI, professeur de microbiologie à l’’IAV
HASSAN II, m’a agréablement accueillie dans son laboratoire. J’ai
pu bénéficier de ses compétences scientifiques et de son savoir faire.
Pour l’accueil chaleureux qu’elle m’a réservé et pour avoir accepté
de faire le rapport de ce mémoire, je lui exprime mes plus vifs
remerciements.
Mes sincères remerciements sont adressés aux professeurs M.
TIJANE et O. BENKHEMMAR pour l’’honneur qu’ils me font de
juger cette thèse.

230
Je désire remercier particulièrement mes collègues les
professeurs A. EL AISSAMI, E. BERRAHO et M. RAHOUTI pour
leur collaboration efficace.
Je désire remercier également Madame AGOUMY, ingénieur
au ministère de l’énergie et des mines pour son aide précieuse lors de
la réalisation de l’analyse minérale des échantillons.
Que mes collègues du laboratoire de Botanique qui m’ont
apportés une aide morale et matérielle trouvent ici l’’expression de
ma meilleure sympathie.
Je remercie vivement M. BENCHAKRI pour son aide
technique efficace et son dévouement.
Enfin, mes remerciements vont à tous les membres de ma
famille pour leur soutien moral et leur encouragement affectueux.

232
Liste des publications
1. REGRAGUI A., LAHLOU H. et ZAID H., 1989. La prémunition de la
tomate contre la verticilliose causée par Verticillium albo-atrum, forme à
microsclérotes. Conséquences physiologiques du phénomène.
Cryptogamie, Mycol. 10 (3) : 243-256
2. REGRAGUI A. ZAID H. and LAHLOU H., 1990. Verticillium albo-
atrum effect on nitrate reductase activity in young tomato plants.
Proceeding of 8th congress of the Mediterranean Phytopathological Union,
Agadir (Morocco) October 28th ,233-235.
3. BALESDENT M.H., REGRAGUI A., LAHLOU H. and BOMPEIX,
1990. Early diagnostic of Verticillium albo-atrum microsclerotia form, and
inoculum evaluation by enzyme linked immunosorbent assay in infected
tomato plants.
Proceeding of 8th congress of the Mediterranean Phytopathological Union,
Agadir (Morocco) October 28th , 75-77.
4. REGRAGUI A., RAHOUTI M. et LAHLOU H., 2003. Effet du stress
salin sur Verticillium albo-atrum: pathogénécité et production d’enzymes
cellulolytiques in vitro
Cryptogamie, Mycol. 24 (2) : 167-174.
5. REGRAGUI A. and LAHLOU H., 2005. Effect of Salinity on in vitro
Trichoderma harzianum Antagonism against Verticillium dahliae.
Pakistan Journal of Biological Sciences 8 (6):872-876
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
1
/
231
100%