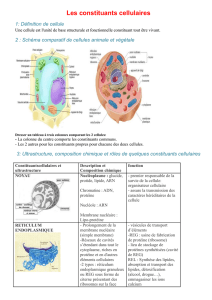
Les polymersomes, des vésicules robustes et

18
Lespolymersomes,
des vésicules robustes
et stimulablespour lalibération
contrôlée desmédicaments
Article proposé par:
ElyesMabrouk,elyes.mabrouk@curie.fr
Damien Cuvelier,damien.cuvelier@curie.fr
FrançoiseBrochard-Wyart,francoise.brochard@curie.fr
Min-HuiLi,min-hui.li@curie.fr
PierreNassoy,pierre.nassoy@curie.fr
LaboratoirePhysico-Chimie Curie, UMR 168, Institut Curie/CNRS/UPMC, Paris
L’utilisation récente de molécules tellesque despolymèrespour formerdes vésiculesaouvert de nouvelles
perspectivesbiomédicales.Ces vésicules, appeléespolymersomes,sontdes sphèrescreusesconstituéesd’une
bicouche de polymères.Dufaitde leur robustesse,lespolymersomes sont utiliséscomme nano-transporteurs de
médicaments destinésàêtre libérésdansles tissus ouorganesmalades.A cesfins thérapeutiques,une étape
importanteconsisteàpouvoirdéclencherladestruction des vésicules une foisla cible atteinte. Nous rapportons
ici desexemplesd’éclatementde polymersomescontrôlé pardes stimuli physiques.Nous discutonsles
mécanismes sous-jacents etleurs applicationspotentielles.
e nombreux médicaments ne peuventpasêtre
administrésdirectementdansl’organisme, carils
sont toxiquespour les tissus sainsou rapidement
éliminésparles reins.Encapsulercesprincipesactifs
dansdes« nano-transporteurs » n’est pasnon plus un
gage absolude protection, caril existe descellules spécia-
lisées(macrophages) dontlafonction est de captureret
d’éliminerlesparticulesétrangèresetpathogènes.Ce pro-
cessus de phagocytose est facilité parl’adsorption de
protéinesàlasurface de laparticule,quisontensuite
reconnues spécifiquementparles récepteurs membra-
nairesdesmacrophages.Pour échapperàlaphagocytose
parlesmacrophages,une desprincipales voies suiviesa
été la conception desnano-transporteurs furtifs,le plus
souventdesliposomesdécoréspardespolymèresde
nature polyéthylèneglycol (PEG). Lesliposomes sontdes
vésiculesdontl’enveloppe est constituée d’une bicouche
de lipidesamphiphiles, c’est-à-dire dont une extrémité est
hydrophile et une terminaison est hydrophobe. Dansle
casde liposomesdécorésde PEG, laprésence de la couche
polymère hydrophile en surface de lavésicule assureune
répulsion d’origine stérique,qui inhibe l’adsorption de
protéinesàleur surface. Toutefois, cesliposomes,méca-
niquementfragiles,ont une durée de vie réduitesous
l’effetducisaillementimposé parle flux sanguin notam-
ment.Récemment,l’utilisation de vésiculespolymères,
baptiséespolymersomes,est apparuecomme une alterna-
tive permettantd’accroîtreletempsde circulation des
nano-transporteurs dansl’organisme. Pour formerces
polymersomes,il suffitd’utiliserdespolymères synthé-
tiquesamphiphiles.Cesmacro-amphiphiles s’associent
alors spontanémenten bicouchesquisereferment sur
elles-mêmespour donnerdes vésicules.La robustesse des
polymersomesaméliorée par rapport aux liposomes vient
essentiellementdufaitque lamasse moléculaire despoly-
mères,qui peut être modulée par synthèsechimique,est
10à100 foisplus élevée quecelle deslipides.L’épaisseur
de la bicouche membranaire peut doncêtreaussi nette-
mentaccrue (voirfigure 1Aetencadré1 ). Cependant,
l’enjeu thérapeutique premiern’est pas uniquement
l’encapsulation de principesactifsdansdesnano-trans-
porteurs ultra-résistants.L’objectif ultime est de faire en
sorte queces réservoirs s’accumulentdansles tissus ou
organesciblésen demeurantintacts aucours de la circula-
tion dansl’organisme,puisde lesdétruire oulesperméa-
biliserpour libérerlesmédicaments une foisarrivésà
destination. Cecahierdeschargesimplique doncune for-
mulation apparemmentantagoniste,puisque les réser-
voirs transporteurs doiventàlafoisêtrerobustesou
furtifs vis-à-visdu système immunitaire etpouvoirêtre
déstabilisésàsouhaitlorsque la cible est atteinte.
Jusqu’àprésent,deux typesde stratégiesontétésui-
viespour réaliser un désassemblage contrôlé de polymer-
somes.La premièrevoie consisteàexploiterles vastes
possibilitésde synthèsechimique pour concevoirdes
membranespolymères sensiblesàl’environnement
chimique. Parexemple,si le caractère hydrophile du
D

19
Lespolymersomes,des vésicules robusteset stimulablespour lalibération contrôlée desmédicaments
copolymèreamphiphile est amplifié en milieuacide,les
polymèresinitialement stablesdansla bicouche sesolubi-
lisentprogressivementdansle milieuaqueux environ-
nant, ce quiconduitàune perméabilisation lente du
polymersome. La deuxième voie,quicommenceàêtre
explorée,viseàutiliserdes stimuli physiques(telsque les
champsélectrique etmagnétique,lalumière oulatem-
pérature) pour détruireàdistancecesnano-réservoirs
polymères.Danslasuite du texte,nous limiteronsnotre
discussion à ceteffetdes stimuli physiques sur les
Figure 1 – (A)Image d’un nano-polymersome obtenue
parcryo-microscopie électroniqueàtransmission. Barre
d’échelle = 20 nm. L’anneau sombre est le cœur hydro-
phobe de la bicouche polymère,d’épaisseur 8 nm (extrait
de B.M.Discheretal.,Science284,1143(1999)).
(B)Imagesd’un polymersome géantobtenue parmicro-
scopie optique,respectivementen fond clair(gauche) et
en fluorescence (droite). Un fluorophore hydrophile est
encapsulé dansle compartimentinterne aqueux dupoly-
mersome. Barre d’échelle = 5 µ m.
Vésiculeslipidiquesetpolymersomes
Encadré 1
Unamphiphile est une molécule constituée d’une partie
hydrophile etd’une partie hydrophobe. Lesdétergents classi-
ques(utilisésdansleslessives) ont une forme conique
(grossetête hydrophile etfine queue hydrophobe) et s’organi-
senten micelles,exposantainsi lapartie hydrophile au
milieuaqueux etécrantantle blochydrophobeaucœur de la
micelle ( figureE1A ). Enrevanche,lorsque laforme molé-
culaire est proche de celle d’uncylindre,lesamphiphilesfor-
ment spontanémentdesbicouches, ce qui permetaux parties
hydrophobes,orientées vers l’intérieur de la bicouche,d’évi-
ter tout contactavecl’eau.Cette forme d’auto-assemblage est
obtenue pour lesphospholipidesqu’on trouve danslesmem-
branesbiologiquesetdontlaqueue hydrophobe est consti-
tuée de deux chaînescarbonées( figureE1B ). Enfin,
lorsqu’une bicouche de moléculesamphiphiles sereferme
sur elle-même,on obtientdes vésiculesdites unilamellaires.
Celles-cisontimperméablesetpeuventêtreutiliséescomme
des réservoirs pour encapsuleretprotégerdesespècesacti-
ves.Troisgrandescatégoriesde vésicules sontactuellement
utiliséesdansle domaine de lalibération contrôlée de
drogues( figureE1C):i) lesliposomes sontconstituésde
phospholipides ; ii) lesliposomesfurtifs sontdesliposomes
«chevelus »,stabilisés stériquement,danslesquelsle phos-
pholipide aété « décoré », auniveaude satête hydrophile,
par une chaîne de polymère hydrosoluble (PEG, polyéthylène
glycol) ;iii) lespolymersomes sontdes vésiculesdansles-
quellesl’amphiphile est un polymère,le plus souvent un
copolymère diblocquicontient unblochydrophile et unbloc
hydrophobe. La possibilité d’utiliserdespolymèresde grande
masse moléculaire (relativementaux phospholipides,et
typiquementde quelques1000 à10 000 g/mol) leur confère
une plus grande épaisseur de membrane,etdoncune imper-
méabilité,une stabilité et une robustesseaccrues vis-à-visdes
contraintesmécaniques.
FigureE1 – Représentation schématique de :(A)une micelle,formée àpartird’amphiphilesde forme conique;(B)une vésicule lipidique, constituée
d’une bicouche de phospholipides ; (C) différentesfamillesde bicouches susceptiblesde formerdes vésicules.Dansle casdupolymersome,le bloc
hydrophile dupolymère joue le rôle de latête polaire dulipide,etleschaîneslipidiquescarbonées sont remplacéesparle blochydrophobe.

Lespolymersomes,des vésicules robusteset stimulablespour lalibération contrôlée desmédicaments
20
vésiculespolymères.Nous nous focaliserons sur deux
approchesdistinctes,quireposent sur desmécanismes
physiques radicalementdifférents.Ilyaquelquesannées,
un groupe américain aétudié laréponse de polymer-
somesàunchamp électrique etmontré que ladestruction
de lamembrane pouvait résulterd’une augmentation de
latension de surface de lavésicule directementinduite
parle champ électrique. Plus récemment, aulaboratoire
de physico-chimie de l’Institut Curie,nous avonsproposé
une stratégie alternative,quireposesur unchangement
de courburespontanée induitparlalumière dansdes
bicouchesasymétriquesde polymèrescristaux liquides.
Avantd’allerplus loin,notonsque,si lesapplications
médicalesdemandentde travailleravecdesnano-liposo-
mesoupolymersomesd’une taille de quelques100 nm,la
plupart des travaux académiques visantà caractériserles
paramètresmécaniquesde cesmembranes sont réalisés
sur des vésiculesditesgéantesd’un diamètre de quelques
10µm (figure 1B );leur visualisation aumicroscope opti-
que permetleur manipulation etleur déformation à
l’échelle de l’objetindividuel. Commençonsdoncpar
introduire quelquesgénéralités sur laphysique desmem-
branesetpar résumerlesprincipalespropriétésméca-
niquesdes vésiculeslipidiquesetpolymères.
Propriétésmécaniques
desmembraneset vésiculesfluides
Aupremierordre,une membrane constituée de deux
feuillets (oumonocouches) de lipidesoude polymères
amphiphiles,secomporte,d’un pointde vue mécanique,
comme une fine tranche homogène,qui peut être
cisaillée, courbée,etcomprimée oudilatée. Al’équilibre,
laréponse de lamembrane à cesdéformationsest caracté-
risée pardesparamètres telsque le module de cisaille-
mentµ,le module de rigidité de courbureκ,etle module
d’étirement surfaciqueK a(quicorrespond àladilatation
oula compression de lamembrane). Les valeurs de ces
modulesdépendentde l’état thermodynamique de la
membrane etde sa composition, c’est-à-dire de lanature
chimique desamphiphilesqui la constituent.Dans un
étatfluide,une membrane n’offreaucune résistanceau
cisaillement,doncµ=0.Lesénergiesélastiquesde cour-
bure etd’étirement sontclassiquementmesuréespar
aspiration de vésiculesdans une micropipette (voirenca-
dré2 ). On définitaussiune tension critiqueàlaquelle la
vésicule éclate:il s’agitde latension de lyseτ c .Aune
échelle moléculaire,lastabilité d’une membrane lipidi-
que oupolymèrerésulte principalementde l’équilibre
entre deux forcesantagonistes:lesforcesattractivesde
Van derWaalsentrechaîneshydrophobesetlesforces
répulsivesde cœur dur (et/ouélectrostatiques) entretêtes
ou segments de chaîneshydrophiles(voirencadré1 ). L’éti-
rementde lamembrane tend àéloignerleslipidesou
chaînesde polymèresles unsdesautres, ce qui expose le
cœur hydrophobe de la bicouche àl’eau, augmenteson
énergie,etcontribueàladéstabilisation de lamembrane.
Cependant,pour atteindreune description réaliste et
complète de laphysique desmembranes,il convientde
prendre explicitementen compte le faitqu’une mem-
brane n’est paspurement un matériaubidimensionnel
élastique ethomogène,maisqu’elle est constituée de
deux feuillets partiellementcouplés.Notamment,l’exis-
tence d’une asymétrie entre lesdeux monocouchespeut
générer une courburespontanée de lamembrane. Cette
asymétrie peut provenir soitd’une différence dansle
nombre de moléculesamphiphilesquicomposentchaque
feuillet,soitd’une différenceauniveaude laforme des
moléculesquiconstituentchaque monocouche. Pour
mieux comprendre l’origine physique de la courbure
spontanée,envisageonsle casconcretd’une membrane
danslaquelle lesdeux feuillets contiennent un nombre
différentd’amphiphiles.Celagénèreune frustration au
sein de lamembrane. Plus précisément,si lasurface est
plane,le feuilletle plus peuplé setrouvecomprimé,tandis
que le feuilletle moinspeuplé est dilaté par rapport àson
étatd’équilibrestable. Il existe doncun gradientde
pression latérale dansladirection de lanormale àlamem-
brane, ce quicrée un momentde flexion. Energéti-
quement,il est alors plus favorable de courberla
membrane pour relâcherla compression dansle feuillet
le plus peuplé etlatension dansle feuilletle moinspeu-
plé. Cettecourburespontanée de lamembrane,C 0 ,est
doncun paramètre intrinsèque de la bicouche,qui
dépend essentiellementde lanaturechimique desespè-
cesamphiphileschoisies.C 0est homogène àl’inverse
d’une longueur,et vaut classiquement une fraction de
nm–1 dansle casde vésiculeslipidiques.En d’autres ter-
mes,la contribution énergétiqueassociée,de l’ordre de
κ C 0 2 ,est généralementnégligeable pour desmembranes
lipidiquesclassiques.
Eclatementde polymersomes
pardesimpulsionsélectriques
L’électroporation est un phénomène largement utilisé
en biologie moléculaire pour incorporer un plasmide (ou
morceaud’ADN) dans une cellule qui permettralafabri-
cation d’une nouvelle protéine,dite exogène,n’existant
pasinitialementdansla cellule. Le principe de base
consisteà appliquer unchamp électriquebref etintense
sur descellules.Cette impulsion électrique provoqueune
perméabilisation transitoire de lamembrane cellulaire et
permetl’entrée dansla cellule de substancesexternes.
Le phénomène d’électroporation,exploité de manière
empirique parlesbiologistes, a été étudié en détail pardes
physiciensen utilisantdesmembranesmodèles,telsque
desliposomesoupolymersomes.Lesmembraneslipidi-
quesoupolymèresétantessentiellementimperméables
aux ions,l’application d’unchamp électrique provoque
l’accumulation deschargesde chaquecôté de la bicouche.
Lecœur hydrophobe de lamembrane,d’épaisseur d hetde
constante diélectriqueε(faible par rapport à celle du

21
Lespolymersomes,des vésicules robusteset stimulablespour lalibération contrôlée desmédicaments
Sonderlespropriétésmécaniquesde vésiculesavecdesmicropipettes
Encadré 2
Lespropriétésmécaniquesdes vésicules sontcaracté-
riséesparplusieurs paramètresassociésaux déformations
appliquéesàla bicouche. La figureE2représentecesdiffé-
rents modesde déformation,àsavoirle cisaillement,la cour-
bure etladilatation. Parmi d’autres,une techniquecourante
qui permetde mesurercertainsde cesparamètresest latech-
nique d’aspiration parmicropipette. Le principe consisteà
aspirer une vésicule individuelle dans une micropipette (de
rayon R p) en créant une dépression àl’aide d’unréservoir
d’eauconnectéàlapipette quise déplaceverticalement
(selon le principe des vasescommunicants). Comme on le
voit sur lafigureE3A ,lavésicule aspirée forme une
« langue » danslapipettetandisque laportion extérieure
demeuresphérique,de rayon R v.Ainsi,latension de la
membrane τ ,qui est homogène àune énergie par unité de
surface,est directementfixée parladépression appliquée
Δ P : selon laloi de Laplace,on a.L’avantage de
cette géométrie réside dansle faitque lalongueur de lalan-
gue, L, donne directementl’augmentation relative d’aire
membranaire pour une tension
donnée. En faisant varierτ ,nous obtenons une relation de
type contrainte-déformation,ouplus précisément tension-
aire de membrane. Généralement,deux régimes sontobser-
vés( figureE3B ). Aux faibles tensions,le régime est ditentro-
pique etl’excèsd’airerécupéré danslapipette provientdes
fluctuationsde lamembrane. Celles-cisontdirectement
reliéesaumodule de rigidité de courbureκde lamembrane,
typiquementde l’ordre de quelquesdizainesde foisl’énergie
thermique (10 –19 J). Augmenterl’aire de lavésicule ( Δ A)a
uncoût énergétiquesimilaireàune tension de surface,que
l’on peut quantifieràl’aide de larelation de Helfrich:
.Aux tensionsplus élevées,lamem-
brane est étirée élastiquementetcaractérisée par un module
d’étirement surfaciqueK a.Ce module caractérise la
compressibilité de la bicouche,et son ordre de grandeur est
de 100 mN/m. Une relation linéaire entretension etdéfor-
mation est alors attendue:.Enfin, aux tensions
encore plus élevées,lavésicule éclate. Cettetension maxi-
male de lyseτ c,de l’ordre de 20 mN/m pour lespolymerso-
mes,est intimementliée àl’épaisseur de lamembrane età
son amincissement sous l’effetd’une déformation dansle
plan de lamembrane. C’est un paramètre qui peut êtreuti-
lisé pour caractériserlarobustesse de lavésicule.
τ≈R·P
pΔ
Δ AA RL R
pv
/02
24≈ππ
FigureE2–Modesde déformation d’une membrane.
ττ πκ
=
00
8
expkT
A
A
B
Δ
τ=KA
A
a
Δ
0
FigureE3–(A)Polymersome géantaspiré dans une micropipette. (B)Courbe de latension de membrane en fonction de ladéformation relative obtenue
parlatechnique de micropipette. En inséré,la courbecomplète permettantde visualiserlatension de lyse.
milieuaqueux environnant)secomporte donc comme
une capacitanceC m(typiquementde l’ordre duµ F/cm 2 ).
Letempsde charge de lamembrane dépend de la conduc-
tivité desmilieux interne etexterne àlavésicule,maisdure
typiquementde l’ordre de lafraction de µsen milieu salin
(5 millimolesparlitre). Par suite,si ladurée de l’impulsion
électrique est bien plus grande que le tempsde charge de
la capacitance,le potentiel transmembranaireV mainsi

Lespolymersomes,des vésicules robusteset stimulablespour lalibération contrôlée desmédicaments
22
généré peut êtreconsidérécomme constantet unique-
mentdépendantduchamp électriqueE appliqué,du rayon
Rde lavésicule etde l’angle θentre ladirection duchamp
etlanormale àlamembrane :V m≈R.E·cos θ .Ce poten-
tiel crée une contrainte électro-compressive perpendicu-
laireauplan de lamembrane, couplée àune tension
latérale . Ceteffet,qui ne modifie pasla
composition desdeux feuillets,donne lieuàune tension
membranaire nette: , comme cela avaitété
introduitparLippmann il yaplus d’unsiècle sous une
forme différentielle. Pour une vésicule polymère initiale-
mentau repos,l’augmentation duchamp électriqueappli-
quésetraduitpar une augmentation de latension nette de
lamembrane. Il existe doncaprioriun potentiel électrique
seuil,V c ,pour lequel latension de lavésicule atteint sa
valeur de lyseτ c .Le groupe de D.Discher, à l’Université de
Pennsylvanie, a testécette prédiction expérimentalement.
Comme on le voit sur lafigure2 ,l’application d’unchamp
électriquecrée deux poresaux pôlesde lavésicule,làoùle
potentiel transmembranaire est maximal( θ=0etπ ).
L’éclaircissementdumilieuinterne de lavésicule vientdu
faitque l’agentde contraste initialementencapsulé (ici,du
sucre) est expulséaucours de l’éclatementde lamem-
brane. Parailleurs,le mécanisme proposé d’augmentation
de tension induite parle champ électriqueaétéconfirmé
quantitativement.Encontrôlantlatension latérale τini-
tiale (c’est-à-dire en l’absence de champ) par une micropi-
pette,il aen effetétévérifié que le potentiel de lyseV c
variaitbien comme ,en accordavecl’équation de
Lippmann.
Alors que l’électroporation est couramment utilisée
pour formerdespores transitoiresdansdescellules sans
quecelane remette en cause leur viabilité,il peut êtresur-
prenantd’observer, comme sur lesphotographiesde la
figure2 ,que l’application d’unchamp électriquesur une
vésicule polymèreconduitàsadésintégration complète.
Cette différence de comportementpeut s’expliquer semi-
quantitativementen regardantde plus prèsle mécanisme
physiqueassocié àlanucléation d’un pore etàsadynami-
que de croissance. Enadaptantàdeux dimensionslathéo-
rie classique de lanucléation de cristaux quireposesur
l’existence de germesen solution,lavariation d’énergie
d’un porederayon rest donnée parlasomme de deux
contributionsénergétiquesantagonistes:d’une part,un
gain en énergie ~–r 2 τ net , associé àlaréduction de surface
membranaire, ce quitend doncàagrandirle pore;d’autre
part uncoût énergétique~+rλ ,oùλest latension de
ligne de lamembrane (voirencadré2 ),quicorrespond à
l’énergie nécessaire pour créerl’interface qui définitle
périmètre du trou, ce quitend àrétrécirle pore. L’énergie
globale dupore présente doncun maximum pour un
rayon critique,typiquementde l’ordre de quel-
quesnanomètresdanslesmembraneslipidiquesoupoly-
mères.Ainsi,lathéorie de lanucléation prévoitquetous
lesporesde taille supérieureà cerayon critiquesont sus-
ceptiblesde croître. Maislasuite duprocessus d’ouver-
ture est un peuplus complexe. Aucours de l’expansion du
pore,latension de membrane relaxe etdufluide interne
est expulsé, carlapression interne,donnée parlaloi de
Laplace,est supérieureàlapression externe àlavésicule.
Deux caspeuventensuitese présenter:i) si le tempsde
fuite duliquide,qui dépend de laviscosité dufluide
interne,est plus petitque le tempscaractéristique
d’ouverture dupore (limité parlaviscosité de lamem-
brane), alors latension de membrane pourraêtrerelaxée
avantdestruction complète;celapermettraune ferme-
ture dupore;ii) àl’inverse,si le fluide interne est vis-
queux,l’expulsion duliquide seraralentie,etle rayon du
pore ne cesserad’augmenter,jusqu’àdésintégration
totale de lavésicule. Dece fait,l’électroporation est une
technique quiconduitàdes résultats trèsdifférents dans
le casdescellulesetdespolymersomes:desporesauto-
cicatrisants sontobservésdanslescellules,sansdoute
parce que lesmembranes sont sous-tenduespar une
«armature » visqueuse (le cytosquelette),etl’électropora-
tion est jugée relativementdoucechezlesbiologistes.Au
contraire dansle casde polymersomesen milieuaqueux,
l’application d’impulsionsélectriquesdétruitlavésicule
de façon irréversible etpermet une soudaine libération à
distance desdroguesencapsulées.Il faut toutefois réaliser
que l’ordre de grandeur duchamp électriqueà appliquer
pour atteindre lalyse est le kV/cm. Appliquerde tels
champsau travers ducorpshumain serait vraisemblable-
mentaccompagné de sévèreseffets secondaires…
Eclatementde polymersomes
parlalumière
Récemment,nous avonsdéveloppé une approche radi-
calementdifférente qui permetd’induire,par unstimulus
physique plus doux,l’éclatementde polymersomes.Pour
ce faire,nous avonsconçudespolymersomes sur mesure,
Figure2–Séquence d’imagesparmicroscopie à contraste de phase de l’écla-
tementd’un polymersome sous l’effetd’unchamp électriqueappliqué dans
ladirection verticale. Lesflèches sur la2 ephotographie montrentlaforma-
tion de deux poresaux deux pôlesde lavésicule. Lesautresdoublesflèches
mettenten évidence la croissance de cespores.Barre d’échelle = 10µ m
(extraitde H.Bermudezetal., Europhys.Lett.64,550(2003)).
σ
el mm
CV=1
22
ττσ
netel
=+
ττ
c−
rnet
*≈λτ
 6
7
8
9
6
7
8
9
1
/
9
100%