CAPES EXTERNE ET CAFEP Section: SCIENCES DE LA VIE ET

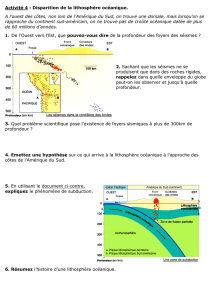
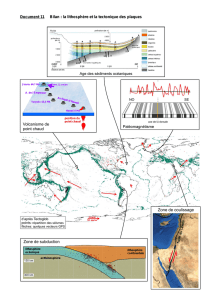
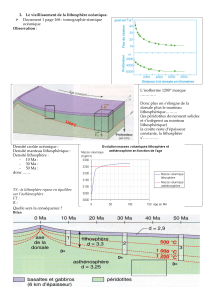
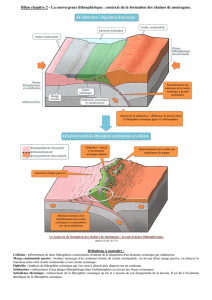
Secrétariat Général
Direction générale des
ressources humaines
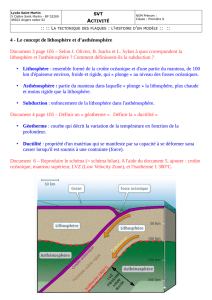
Sous-direction du recrutement
Concours du second degré – Rapport intermédiaire du jury
Session 2014 exceptionnelle
CAPES EXTERNE ET CAFEP
Section: SCIENCES DE LA VIE ET DE LA TERRE
Rapport de jury présenté par Gilles MERZERAUD
Président de jury
Les rapports des jurys des concours sont établis sous la responsabilité des présidents
de jury

SOMMAIRE
- Introduction et statistiques.
- Sujet d’écrit de Biologie.
- Correction du sujet de Biologie et conseils aux candidats.
- Sujet d’écrit de Géologie.
- Correction du sujet de Géologie et conseils aux candidats.

CAPES CAFEP
Biologie Géologie Biologie Géologie Biologie Géologie Biologie Géologie Biologie Géologie Biologie Géologie
Note Min, 3,19 3,19 4,5 4,55 3,12 2,45 5,1 2,8 4,16 2,3 5,81 2,95
Note Max, 16,31 19,2 15,19 18,6 17,8 17 15,1 15,1 18,41 13,91 15,63 12,03
Ecart-type 2,23 2,2 2,53 2,3 3,5 3,6 2,9 3,3 2,66 2,17 2,05 1,83
Moyenne des
admissibles
8,52 9,64 8,78 9,64 11,2 9,2 10,3 8,7 11,45 8,13 10,39 7,2
CAPES CAFEP
2013-2
ADMISSIBILITE sur 3 ans
2012
CAPES CAFEP
2013
Introduction et statistiques
Pour la session 2014 exceptionnelle, que nous appellerons ici 2013 – (2), 420 postent étaient ouverts au
concours pour le CAPES et 85 pour le CAFEP.
Le taux d’admissibilité a été de 1.07 x le nombre de postes et donc 692 candidats ont été reconnus
admissibles au CAPES et 92 au CAFEP.
Ces chiffres incluent les candidats déjà admis en 2013 au CAPES-CAFEP SVT ainsi que ceux admis à
l’agrégation SV-STU. Sur 2862 candidats 1737 se sont présentés à l’oral n°1 (soit 61%) et 1676 se sont
présentés à l’Oral n°2 (soit 58%).
Statistiques des résultats d’admissibilité comparées entre la session 2013 et 2013(2)
2013 -2 2013
2012

Bilan d’admission comparés sur les 3 dernières années.
La brusque rupture entre les sessions 2013 et 2013 (2) est liée au fait qu’aucune correction des notes n’a
été réalisée pour remonter les notes, du fait du mode de correction en chapelle et des délais de
correction. On constatera par ailleurs une baisse irrégulière mais sensible du niveau scientifique des
candidats depuis 3 ans.
Moyenne des candidats non éliminés
Moyenne des admissibles
Barres d’admissibilité

Sujet d’écrit de Biologie
SESSION 2014 anticipée
CAPES
CONCOURS EXTERNE
ET CAFEP
Section : SCIENCES DE LA VIE ET DE LA TERRE
COMPOSITION SUR UN SUJET DE BIOLOGIE
Durée : 5 heures
L‘usage de tout ouvrage de référence, de tout dictionnaire et de tout matériel électronique est
rigoureusement interdit.
Dans le cas où un(e) candidat (e) repère ce qui lui semble être une erreur d’énoncé, il (elle) le signale
très lisiblement sur sa copie, propose la correction et poursuit l’épreuve en conséquence. De même, si
cela vous conduit à formuler une ou des hypothèses, il vous est demandé de la (ou les) mentionner
explicitement.
NB :
- Hormis l’en-tête détachable, la copie que vous rendrez devra, conformément au principe d’anonymat,
ne comporter aucun signe distinctif, tel que nom, signature, origine, etc.
- Si le travail qui vous est demandé comporte notamment la rédaction d’un projet ou d’une note, vous
devrez impérativement vous abstenir de signer ou de l’identifier.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
1
/
70
100%