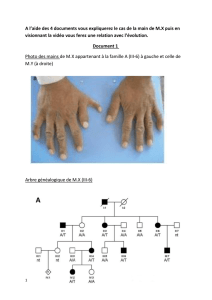
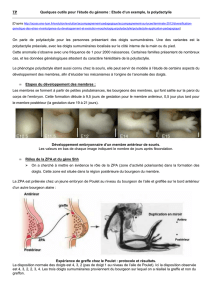
MR61431

Universite de Sherbrooke
CARACTERISATION DE LA SIGNALISATION ET DES ROLES FONCTIONNELS
DES HEDGEHOGS DANS LA MORPHOGENESE GASTROINTESTINALE
Par:
Sabrina Turgeon
Departement d'anatomie et de biologie cellulaire
Memoire presente a la faculte de medecine et des sciences de la sante
en vue de l'obtention du grade de maitre es sciences (M.Sc.) en biologie cellulaire
Sherbrooke, Quebec, Canada, 24 Aout 2009
Membre du jury:
Professeur
:
Daniel Menard, programme de biologie cellulaire
Professeure : Nathalie Perreault, programme de biologie cellulaire
Professeure Marie-Josee Boucher, programme de biologie cellulaire
Professeur Louis Gendron, programme de physiologie et de biophysique

1*1 Library and Archives
Canada
Published Heritage
Branch
395 Wellington Street
OttawaONK1A0N4
Canada
Bibliotheque et
Archives Canada
Direction du
Patrimoine de
I'edition
395,
rue Wellington
OttawaONK1A0N4
Canada
Your file Voire reference
ISBN: 978-0-494-61431-0
Our file Notre r6f6rence
ISBN: 978-0-494-61431-0
NOTICE:
AVIS:
The author has granted a
non-
exclusive license allowing Library and
Archives Canada to reproduce,
publish,
archive, preserve, conserve,
communicate to the public by
telecommunication or on the Internet,
loan,
distribute and sell theses
worldwide, for commercial or
non-
commercial purposes, in microform,
paper, electronic and/or any other
formats.
L'auteur
a accorde une licence non exclusive
permettant a la Bibliotheque et Archives
Canada de reproduire, publier, archiver,
sauvegarder, conserver, transmettre au public
par telecommunication ou par
I'lnternet,
preter,
distribuer et vendre des theses partout dans le
monde, a des fins commerciales ou autres, sur
support microforme, papier, electronique et/ou
autres formats.
The author retains copyright
ownership and moral rights in this
thesis. Neither the thesis nor
substantial extracts from it may be
printed or otherwise reproduced
without the author's permission.
L'auteur
conserve la propriete du droit d'auteur
et des droits moraux qui protege cette these. Ni
la these ni des extra its substantiels de celle-ci
ne doivent etre imprimes ou autrement
reproduits sans son autorisation.
In compliance with the Canadian
Privacy Act some supporting forms
may have been removed from this
thesis.
Conformement a la loi canadienne sur la
protection de la vie privee, quelques
formulaires secondaires ont ete enleves de
cette these.
While these forms may be included
in the document page count, their
removal does not represent any loss
of content from the thesis.
Bien que ces formulaires aient inclus dans
la pagination, il n'y aura aucun contenu
manquant.
1*1
Canada

CARACTERISATION DE LA SIGNALISATION ET DES ROLES
FONCTIONNELS DES HEDGEHOGS DANS LA MORPHOGENESE
GASTROINTESTINALE
Par : Sabrina Turgeon
Memoire presente a la Faculte de Medecine et des Sciences de la Sante en vue de
l'obtention du diplome de maitre es sciences, Departement d'anatomie et de biologie
cellulaire, Faculte de Medecine et des Sciences de la Sante, Universite de Sherbrooke,
Sherbrooke, Quebec, Canada, J1H 5N4
RESUME
Les Hedgehogs (Hh) sont des morphogenes multifonctionnels qui ont des roles importants
dans l'embryogenese et la morphogenese de plusieurs organes. Leur role dans le
developpement de la glande gastrique humaine n'est pas connu. Peu est connu de la
signalisation Hh, plus precisement de Sonic Hedgehog (Shh), dans la glande gastrique
humaine adulte, autant dans sa localisation et son association aux differents types
cellulaires gastriques que dans son role fonctionnel dans cet organe. Les modeles murins
de deletion classique des molecules de la signalisation Hh subissent, dans la grande
majorite, une mort neonatale ou au stade somites rendant ces modeles inutiles pour
l'etude du maintien de l'homeostasie du tube digestif adulte, autant au niveau de l'intestin
que de l'estomac. Dans un premier temps, les patrons d'expression des differents genes
cles impliques dans la voie de signalisation Hh dans le developpement foetal humain de la
glande gastrique ont ete determines a l'aide de tissus gastriques fcetaux ages entre 14 et 20
semaines de gestation. Dans un deuxieme temps, les roles fonctionnels et physiologiques
de Shh, dans l'homeostasie de l'intestin adulte et dans la morphogenese et homeostasie de
l'estomac ont ete elucides par la generation de souris possedant une deletion
conditionnelle de Shh a 1'epithelium de l'intestin ou de l'estomac a l'aide du systeme
Cre/loxP.
L'analyse des patrons d'expression des ligands SHH et Indian Hedgehog (IHH), des
recepteurs PATCHED et SMOOTENED, des effecteurs GLI et de genes cibles de la voie
de signalisation Hh dans la glande gastrique humaine en developpement, a confirme la
presence de la majorite de ces proteines des 14 semaines de gestation. Ces effecteurs
n'ont ete detectes qu'au niveau des cellules a mucus de surface, des cellules du foveolae
et des cellules zymogeniques mais pas au niveau des cellules parietales. L'expression
strictement epitheliale de presque toutes ces proteines suggere fortement
1'activation
exclusivement autocrine de cette voie de signalisation dans la glande gastrique humaine
foetale. Ces patrons d'expression different de ceux retrouves dans la glande gastrique
murine foetale ou adulte.
L'etude du modele murin V////«e-Cre;SMIoxP/loxP, qui possede une deletion de Shh a
l'epithelium intestinal, a permis d'elucider les roles fonctionnels de Shh dans
l'homeostasie de l'intestin. Nous avons constate que Shh a un role specifique dans
l'homeostasie intestinale puisque les souris experimentales presentent un allongement

villositaire. Shh est egalement implique dans la maturation des cellules caliciformes et
dans l'inhibition de la proliferation epitheliale intestinale. Les resultats montrent aussi
que Ihh semble, quant a lui, avoir un role compensateur puisqu'il se relocalise dans les
regions qui expriment normalement Shh.
L'etude du modele murin Foxa3-Cre;ShhloxPnoxP, qui possede une deletion de Shh dans
1'epithelium gastrique, a permis d'elucider les roles de Shh dans le developpement et le
maintien de la glande gastrique. Nous avons determine que Shh a un role dans la
morphogenese de la glande gastrique et dans le maintien de ce dernier. En effet,
1'architecture foveolae-glande est fortement affectee par la perte epitheliale de Shh. Les
resultats demontrent egalement que Shh a un role determinant dans la differentiation des
cellules parietales puisque ces dernieres sont absentes chez les souris experimentales.
Mots cles : Shh et Ihh, Cellules epitheliales intestinales et gastriques, Facteur de
transcription Gli, Bmp, Morphogenese.

A la memoire de mon
grand-pere Jean-Claude
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
1
/
135
100%