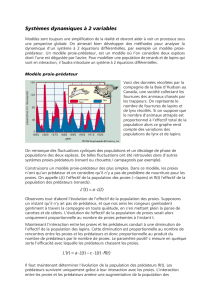
CAPES Rappels Dymanique des Pop et Ecologie

Populations, peuplements et communautés
-Croissance et dynamique des populations
-Interactions entre populations : compétition interspécifiques, niche écologique,
relations prédateurs -proies et hôtes -parasites, approche qualitative du
formalisme de Lotka-Volterra, conception évolutive des interactions durables
-Notion de peuplement (guildes), succession écologique (peuplements pionniers et
climaciques, zonation)
Écologie fonctionnelle, écosystèmes
-Zonation écologique au niveau de la planète (biomes terrestres et océaniques)
-Le sol et ses composants
-Flux d'énergie, réseaux et pyramides trophiques, étude d'un écosystème et
comparaison avec un agrosystème
-Participation des êtres vivants aux cycles de l'eau, du carbone et de l'azote,

Biodiversitéet biologie de la conservation
-Définition et portée du concept de biodiversité. Échelles (génétique,
spécifique, écologique)
-Dynamique de la biodiversité(maintien, extinction d'origine anthropique)
-Biologie de la conservation et gestion durable des populations

Un nénuphar, Victoria regia, double de surface tous les 9 jours. Après 3 mois, un seul
nénuphar a couvert entièrement la moitiéd’une pièce d’eau. Combien faudra-t-il de
temps pour couvrir l’autre moitié?

Croissance d’une population
N0= 1
N1= R.N0
N2= R.N1= R.R.N0 = R2.N0
N3= R.N2= R.R.R.N0 = R3.N0
N4= R.N3= R.R.R.R.N0 = R4.N0
N5= R.N4= R.R.R.R.R.N0 = R5.N0
……
Nt= Rt.N0
0
5000
10000
0 5 10 15 20 25 30
Temps
Effectif
c’est une croissance exponentielle
Nt= ert.N0
avec R=er

on a :
N N ert
=
0
Cela revient àdire qu’en l’absence de facteur limitant,
une population s’accroît de façon exponentielle
en posant N0= 1, on a :
N
e
rt
=
en se rappelant que dN
dt NdN
dt
(ln )
=
1
ou :
ln
N
rt
=
1
NdN
dt r
=
ou :
dN
dt rN
=
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
1
/
86
100%