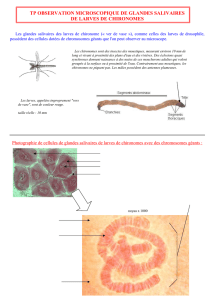
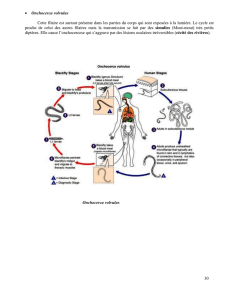
contribution bibliographique à l`étude des produits d

ANNEE 2005 THESE : 2005 – TOU 3 – 4075
CONTRIBUTION BIBLIOGRAPHIQUE
À L’ÉTUDE DES PRODUITS
D’EXCRÉTION/SÉCRÉTION
DES LARVES D’OESTRE ET APPARENTÉS
_________________
THESE
pour obtenir le grade de
DOCTEUR VETERINAIRE
DIPLOME D’ETAT
présentée et soutenue publiquement en 2005
devant l’Université Paul-Sabatier de Toulouse
par
Jérôme, Pascal FOSTINELLI
Né, le 18 avril 1979 à LIBOURNE (Gironde)
___________
Directeur de thèse : Mme le Docteur Lydie BRET-BENNIS
___________
JURY
PRESIDENT :
M. Alexis VALENTIN
ASSESSEUR :
Mme Lydie BRET-BENNIS
M. Philippe JACQUIET
Professeur à l’Université Paul-Sabatier de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE

Toulouse, 2005
NOM : FOSTINELLI PRENOM : Jérôme Pascal
TITRE : CONTRIBUTION A L’ETUDE BIBLIOGRAPHIQUE DES PRODUITS
D’EXCRETION / SECRETION DES LARVES D’OESTRE ET APPARENTES
RESUME :
Les insectes Diptères Oestrus ovis, Hypoderma lineatum et Lucilia cuprina sont
responsables de myiases chez les ruminants engendrant d’importantes pertes
économiques. De nouvelles stratégies de lutte sont à l’étude, notamment celles
basées sur le développement de vaccins à base d’antigènes extraits des produits
d’excrétion/sécrétion des larves. Ces derniers sont majoritairement des sérine-
protéases d’origine digestive ayant des activités enzymatiques similaires d’une
espèce larvaire à l’autre. Ces protéases contribuent au développement des larves :
elles assurent leur nutrition, leur établissement, leur migration et leur protection vis-à-
vis du système immunitaire de l’hôte. Les tentatives de vaccination utilisant des
antigènes extraits des produits d’excrétion/sécrétion se sont soldées par un échec.
Cependant, plusieurs pistes restent à explorer notamment chez Oestrus ovis.
MOTS CLES :
Oestrus ovis, Hypoderma lineatum, Lucilia cuprina, produits d’excrétion/sécrétion,
myiase, larve.
TITLE: BIBLIOGRAPHIC CONTRIBUTION TO THE STUDY OF EXCRETION /
SECRETION PRODUCTS OF OESTRE LARVAE AND RELATED.
ABSTRACT:
The Diptera Oestrus ovis, Hypoderma lineatum and Lucilia cuprina are responsible
for myiasis in ruminants causing significant economics losses. New counter
strategies are being studied, one in particular concerning the development of
vaccines using the antigen base extracts from larval excretion/secretion products.
These are mainly serine-protease developed during digestion and have an enzymatic
behaviour similar from one larval species to the next.
These proteases contribute to the larval development and ensure their nutrition,
migration and protection from the host immune system.
Vaccination attempts using antigen extracts from larval excretion/secretion products
result in failure. Nonetheless, several leads remain to be studied, especially in the
case of Oestrus ovis.
KEYWORDS:
Oestrus ovis, Hypoderma lineatum, Lucilia cuprina, excretion/secretion products,
myiasis, larvae.


MINISTERE DE L'AGRICULTURE ET DE LA PECHE
ECOLE NATIONALE VETERINAIRE DE TOULOUSE
Directeur : M. P. DESNOYERS
Directeurs honoraires……. : M. R. FLORIO
M. J. FERNEY
M. G. VAN HAVERBEKE
Professeurs honoraires….. : M. A. BRIZARD
M. L. FALIU
M. C. LABIE
M. C. PAVAUX
M. F. LESCURE
M. A. RICO
M. A. CAZIEUX
Mme V. BURGAT
M. D. GRIESS
M. J. CHANTAL
M. J.-F. GUELFI
M. M. EECKHOUTTE
PROFESSEURS CLASSE EXCEPTIONNELLE
M. CABANIE Paul, Histologie, Anatomie pathologique
M. DARRE Roland, Productions animales
M. DORCHIES Philippe, Parasitologie et Maladies Parasitaires
M. TOUTAIN Pierre-Louis, Physiologie et Thérapeutique
PROFESSEURS 1ère CLASSE
M. AUTEFAGE André, Pathologie chirurgicale
M. BODIN ROZAT DE MANDRES NEGRE Guy, Pathologie générale, Microbiologie, Immunologie
M. BRAUN Jean-Pierre, Physique et Chimie biologiques et médicales
M. DELVERDIER Maxence, Histologie, Anatomie pathologique
M. EUZEBY Jean, Pathologie générale, Microbiologie, Immunologie
M. FRANC Michel, Parasitologie et Maladies Parasitaires
M. MARTINEAU Guy-Pierre, Pathologie médicale du Bétail et des Animaux de basse-cour
M. MILON Alain, Pathologie générale, Microbiologie, Immunologie
M. PETIT Claude, Pharmacie et Toxicologie
M. REGNIER Alain, Physiopathologie oculaire
M. SAUTET Jean, Anatomie
M. SCHELCHER François, Pathologie médicale du Bétail et des Animaux de basse-cour
PROFESSEURS 2e CLASSE
Mme BENARD Geneviève, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
M. BERTHELOT Xavier, Pathologie de la Reproduction
M. CONCORDET Didier, Mathématiques, Statistiques, Modélisation
M. CORPET Denis, Science de l'Aliment et Technologies dans les industries agro-alimentaires
M. DUCOS Alain, Zootechnie
M. DUCOS DE LAHITTE Jacques, Parasitologie et Maladies parasitaires
M. ENJALBERT Francis, Alimentation
M. GUERRE Philippe, Pharmacie et Toxicologie
Mme KOLF-CLAUW Martine, Pharmacie -Toxicologie
M. LEFEBVRE Hervé, Physiologie et Thérapeutique
M. LIGNEREUX Yves, Anatomie
M. PICAVET Dominique, Pathologie infectieuse
PROFESSEUR ASSOCIE
M. HENROTEAUX Marc, Médecine des carnivores
INGENIEUR DE RECHERCHES
M. TAMZALI Youssef, Responsable Clinique équine
PROFESSEURS CERTIFIES DE L’ENSEIGNEMENT AGRICOLE
Mme MICHAUD Françoise, Professeur d'Anglais
M. SEVERAC Benoît, Professeur d'Anglais

MAÎTRE DE CONFERENCES HORS CLASSE
M. JOUGLAR Jean-Yves, Pathologie médicale du Bétail et des Animaux de basse-cour
MAÎTRE DE CONFERENCES CLASSE NORMALE
M. ASIMUS Erik, Pathologie chirurgicale
M. BAILLY Jean-Denis, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
M. BERGONIER Dominique, Pathologie de la Reproduction
M. BERTAGNOLI Stéphane, Pathologie infectieuse
Mme BOUCRAUT-BARALON Corine, Pathologie infectieuse
Mlle BOULLIER Séverine, Immunologie générale et médicale
Mme BOURGES-ABELLA Nathalie, Histologie, Anatomie pathologique
M. BOUSQUET-MELOU Alain, Physiologie et Thérapeutique
Mme BRET-BENNIS Lydie, Physique et Chimie biologiques et médicales
M. BRUGERE Hubert, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
Mlle CADIERGUES Marie-Christine, Dermatologie
Mme CAMUS-BOUCLAINVILLE Christelle, Biologie cellulaire et moléculaire
Mme COLLARD-MEYNAUD Patricia, Pathologie chirurgicale
Mlle DIQUELOU Armelle, Pathologie médicale des Equidés et des Carnivores
M. DOSSIN Olivier, Pathologie médicale des Equidés et des Carnivores
M. FOUCRAS Gilles, Pathologie du bétail
Mme GAYRARD-TROY Véronique, Physiologie de la Reproduction, Endocrinologie
M. GUERIN Jean-Luc, Productions animales
Mme HAGEN-PICARD Nicole, Pathologie de la Reproduction
M. JACQUIET Philippe, Parasitologie et Maladies Parasitaires
M. JAEG Jean-Philippe, Pharmacie et Toxicologie
M. LYAZRHI Faouzi, Statistiques biologiques et Mathématiques
M. MARENDA Marc, Pathologie de la reproduction
M. MATHON Didier, Pathologie chirurgicale
Mme MESSUD-PETIT Frédérique, Pathologie infectieuse
M. MEYER Gilles, Pathologie des ruminants
M. MONNEREAU Laurent, Anatomie, Embryologie
Mme PRIYMENKO Nathalie, Alimentation
Mme RAYMOND-LETRON Isabelle, Anatomie pathologique
M. SANS Pierre, Productions animales
Mlle TRUMEL Catherine, Pathologie médicale des Equidés et Carnivores
M. VERWAERDE Patrick, Anesthésie, Réanimation
MAÎTRE DE CONFERENCES CONTRACTUELS
M. CASSARD Hervé, Pathologie du bétail
N. DESMAIZIERES Louis-Marie, Clinique équine
M. LEON Olivier, Elevage et santé en productions avicoles et porcines
MAÎTRE DE CONFERENCES ASSOCIE
M. REYNOLDS Brice, Pathologie médicale des Equidés et Carnivores
ASSISTANTS D'ENSEIGNEMENT ET DE RECHERCHE CONTRACTUELS
M. CORBIERE Fabien, Pathologie des ruminants
Mlle LACROUX Caroline, Anatomie pathologique des animaux de rente
Mme MEYNADIER-TROEGELER Annabelle, Alimentation
M. MOGICATO Giovanni, Anatomie, Imagerie médicale
Mlle PALIERNE Sophie, Chirurgie des animaux de compagnie
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
1
/
105
100%