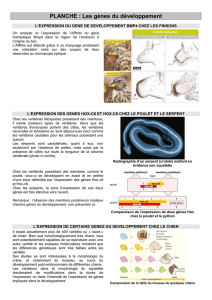
THÈSE Découverte de biomarqueurs prédictifs en cancer du sein

Aix-Marseille Université
Faculté des Sciences de Luminy
École Doctorale des Sciences de la Vie et de la Santé
Noattribué par la bibliothèque
|_|_|_|_|_|_|_|_|_|_|_|_|
THÈSE
Présentée en première version
le 18 octobre 2013 par
Maxime Ulysse Garcia
Né le 14 mai 1982 à Arles (13)
Découverte de biomarqueurs
prédictifs en cancer du sein par
Intégration
Transcriptome-Interactome
Pour l’obtention du grade de
Docteur d’Aix-Marseille Université
Spécialité : Bioinformatique
Jury :
M. JACQUES VAN HELDEN AMU - TAGC (Président)
M. PHILIPPE DESSEN Génétiques des Tumeurs (Rapporteur)
M. BENNO SCHWIKOWSKI Institut Pasteur (Rapporteur)
M. PASCAL BARBRY IPMC (Examinateur)
M. FRANÇOIS BERTUCCI IPC (Directeur)
M. GHISLAIN BIDAUT AMU - CRCM (Co-directeur)


À ma famille
À mes parents
À Célia

REMERCIEMENTS
Je voudrais tout d’abord exprimer mes plus profonds remerciements à Ghislain sans qui
cette thèse n’aurait pas eu lieu, et qui en plus d’un encadrement sans faille depuis le début
a été une source de conseils tout au long de cette thèse. Je remercie de même François
pour toute l’aide qu’il a pu m’apporter, pour la rapidité, la justesse de ses réponses et ses
corrections à toutes heures. Je voudrais également remercier pour les conseils qu’elles m’ont
apportées, toutes les personnes avec qui j’ai collaboré pour cette thèse : Daniel, Pascal,
Arnaud, Sabrina, Raphaële, Renaud.
Je tiens également à exprimer mes plus sincères remerciements à tous les membres de la
plateforme de Bioinformatique Intégrative du
CRCM
qui m’ont supporté malgré eux tout le
long de ma thèse et de mon écriture : Samuel, Olivier, Alexandre, Fanny, Claire, Quentin,
Guillaume. Je voudrais également inclure dans ces remerciements l’ensemble du CRCM :
Françoise et Jean-Paul qui ont soutenu ce projet dès le début, Caroline pour avoir été un
soutien moral sans faille, Sébastien, Will, Sébastien, Marie-José, Yves, Khedidja, Avais, MLA,
MLT, Vincent, Javier, Émilie, Marion, Julien, Armelle, Axelle, Pascale, Bernard, Myriam,
Julie, François, Laurence D, Laurence L et je présente mes excuses à tous ceux que j’aurais
très probablement oublié...
Je souhaite aussi adresser mes remerciements à tous mes amis qui m’ont soutenu ces
dernières années, mais qui sont trop nombreux pour les citer tous ici.. .
J’exprime toute ma gratitude à mes parents Claude et Marie-Claude, mon frère Florent,
Karine et leur trois enfants Robin, Thibaud et Tristan ainsi que tout le reste de la famille.. .
Je conclurai en remerciant de tout mon cœur Célia pour son aide et son soutien.
iv

TABLE DES MATIÈRES
Thèse i
Table des Matières v
Index des Tableaux viii
Index des Figures ix
I Introduction générale 2
I.1 Problématique ..................................... 3
I.2 La recherche sur le cancer .............................. 5
I.2.a Historique ................................... 5
I.2.b Rappels sur l’expression des gènes et sa régulation ........... 7
I.2.c Les caractéristiques spécifiques des cancers ................ 13
I.3 L’apport des technologies à haut-débit à la recherche sur le cancer . . . . . . . 22
I.3.a L’ère post-génomique et la fin du paradigme un gène, une maladie . . . 22
I.3.b La médecine prédictive et la médecine personnalisée . . . . . . . . . . 24
I.4 La recherche sur le cancer du sein .......................... 26
I.4.a Les caractéristiques du cancer du sein ................... 26
I.4.b Diagnostics et traitements .......................... 28
I.4.c Les classifications utilisées dans le cancer du sein ............ 30
I.4.d Intérêts des signatures prédictives dans le cancer du sein . . . . . . . . 34
I.4.e Les limitations des technologies utilisées ................. 37
I.4.f Les solutions ................................. 38
I.5 Conclusion ....................................... 40
v
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
1
/
201
100%