Cours

Chapitre 11 – L'immunité adaptative
Les bactéries sont des cellules (procaryotes, sans noyau) qui se multiplient par des divisions
cellulaires successives. La plupart des bactéries pathogènes se multiplient dans le milieu extra-
cellulaire. (Exceptions : Listeria, et aussi le bacille de Koch, responsable de la tuberculose, qui se
multiplie dans les cellules pulmonaires.)
Les virus, en revanche, ne sont pas des cellules et ont obligatoirement besoin, pour se multiplier,
de pénétrer à l'intérieur d'une cellule de notre organisme et de détourner son fonctionnement
pour lui faire fabriquer de nouvelles particules virales.
Comment l'immunité adaptative permet-elle d'éliminer efficacement les pathogènes présents dans le
milieu extra-cellulaire ? Même question pour les cellules infectées par un pathogène (ou devenues
anormales, ex. cancéreuses).
I. L'immunité adaptative est spécifique
Voir activité 24 et doc. 1 p. 310. Tétanos et diphtérie sont deux maladies différentes dues à des
bacilles (=bactéries en forme de bâtonnets) : bacille tétanique et bacille diphtérique. Ces bacilles se
multiplient dans le milieu extra-cellulaire de l'organisme ; chacun produit une toxine, petite protéine
toxique pour l'organisme, qui est responsable des symptômes de la maladie.
La toxine tétanique entraîne le blocage en position contractée (=tétanos) des muscles ; les
symptômes commencent autour de l'endroit où les bacilles ont pénétré, puis se propagent à tout
l'organisme ; la mort survient par blocage des muscles de la cage thoracique normalement
responsables des mouvements respiratoires.
La toxine diphtérique entraîne les symptômes d'une très grave angine et aboutit à la mort du patient.
Le vaccin anti-tétanique consiste en l'injection de la toxine tétanique partiellement dénaturée par
chauffage. D'après le doc. 1 p. 310, on constate que le vaccin contre un agent pathogène donné
(ici le bacille tétanique) prépare le système immunitaire à neutraliser rapidement et efficacement
ce pathogène (ce qui permet la survie des cobayes). Cette réponse, dite adaptative parce
qu'élaborée suite à une première rencontre avec le pathogène, est spécifique : elle ne protège pas
contre un autre agent pathogène (le bacille diphtérique ici).
II. L'immunité adaptative implique différentes catégories de lymphocytes, qui coopèrent
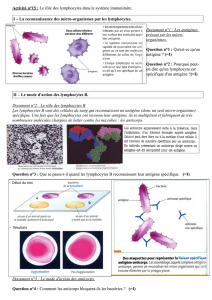
Doc. 3 p. 311. Les lymphocytes font partie des leucocytes (globules blancs) ; ce sont des
cellules grossièrement sphériques, pourvues d'un gros noyau. Différentes catégories de
lymphocytes se distinguent par les molécules qu'ils portent à leur surface :
– lymphocytes B : présence de …………………… (= anticorps membranaires) à leur surface;
– lymphocytes T : présence de ……………………… à leur surface, avec en plus :
– la protéine …………… à la surface des lymphocytes T CD4 (ou "T4") ;
– la protéine …………… à la surface des lymphocytes T CD8.
Lire le document 2 p. 310. En déduire quel est le principal agent par lequel l'immunité adaptative aide
à éliminer les antigènes extracellulaires (toxine diphtérique ici). Même question pour les antigènes
intracellulaires (bacille de Koch ici). Justifier.
……………………………………………………………………………………………………………………
……………………………………………………………………………………………………………………
……………………………………………………………………………………………………………………
……………………………………………………………………………………………………………………
Doc. 4 p. 311. On prélève les lymphocytes de souris en draînant leur circulation lymphatique à l'aide
d'un tuyau, et on place ces lymphocytes en culture. Ensuite, on arrête la production de cellules
immunitaires chez ces souris par irradiation. On réinjecte alors à ces souris seulement certaines
catégories de lymphocytes. Que peut-on déduire des résultats de cette expérience ?
……………………………………………………………………………………………………………………
……………………………………………………………………………………………………………………
Chapitre 11 – L'immunité adaptative 1 / 8

D'après cette expérience, quelles cellules sécrètent les anticorps ? Qu'est-ce qui
permet de l'affirmer ?
……………………………………………………………………………………………
……………………………………………………………………………………………
……………………………………………………………………………………………
……………………………………………………………………………………………
……………………………………………………………………………………………
…………………………………………………………………………………………………………………………………………………………………………
…………………………………………………………………………………………………………………………………………………………………………
2
lymphocytes
globules
rouges de
mouton
(GRM)

Tous les lymphocytes sont produits au départ dans la moelle osseuse rouge (doc. 3 p. 325).
- Les lymphocytes B (B pour Bone marrow : moelle osseuse) subissent leur maturation directement
dans la moelle osseuse ;
- tandis que les lymphocytes T (T pour thymus), après avoir quitté la moelle osseuse, doivent aller
dans le thymus, organe situé en arrière du sternum, pour subir une maturation. Voir paragraphe VII.
Moelle osseuse et thymus sont les organes lymphoïdes primaires. Ces organes libèrent des
lymphocytes B et T naïfs (=n'ayant pas encore rencontré l'antigène contre lequel ils sont
spécifiquement dirigés). Ces lymphocytes naïfs circulent ensuite régulièrement d'un organe
lymphoïde secondaire (ganglions lymphatiques...) à un autre en empruntant la circulation
lymphatique et la circulation sanguine. S'ils rencontrent l'antigène qu'ils sont capables de
reconnaître spécifiquement, ils se multiplient alors et se différencient (voir III, IV, V).
III. Les lymphocytes T auxiliaires, pivots des réactions immunitaires adaptatives
1. Activation des lymphocytes T CD4 en LT auxiliaires
On s'intéresse ici aux lymphocytes T CD4. (Le mécanisme d'activation des LT CD8 est semblable,
mais le résultat est différent, voir paragraphe V.)
Doc. 3 p. 313. Les lymphocytes T portent des récepteurs T qui reconnaissent spécifiquement
l'antigène (sous la forme d'un fragment peptidique issu de la digestion du pathogène lors de la
phagocytose) associé à une molécule du CMH, le tout porté à sa surface par une cellule
présentatrice de l'antigène (voir chapitre 10 – II. 1).
Seuls les lymphocytes portant à leur surface un récepteur T de forme complémentaire de celle de
l'antigène vont alors réagir : on parle de sélection clonale. Ces lymphocytes spécifiques de l'antigène
vont alors se multiplier, formant en quelques jours un clone de milliers de cellules ( prolifération). La
plupart des cellules du clone vont alors se différencier en lymphocytes T auxiliaires (ou LTH,
H=helper).
Cependant, certaines cellules du clone resteront provisoirement inactives et ne s'activeront que lors
de la prochaine rencontre avec le même antigène : ce sont des cellules mémoire.
2. Rôle des LT auxiliaires
Les lymphocytes T auxiliaires sécrètent des cytokines (dont l'interleukine 2 ou IL2 qui stimule la
prolifération des LT CD4 et CD8 activés). Certaines cytokines stimulent les cellules
phagocytaires sur le lieu de l'infection (voir chapitre 10-II.2). Elles stimulent également l'activation
des lymphocytes B spécifiques du même antigène (voir IV.4).
IV. Les anticorps, agents du maintien de l’intégrité du milieu extracellulaire
1. Reconnaissance de l'antigène par les immunoglobulines
Doc. 1 et 2 p. 312 + activité 25. Toutes les
immunoglobulines (=anticorps) sont
constituées de quatre chaînes, deux
chaînes lourdes et deux chaînes légères.
Chaque chaîne comporte une partie
constante et une partie variable. La partie
variable des chaînes lourdes et légères
est responsable de la reconnaissance
spécifique de l'antigène.
Les deux sites de reconnaissance d'une
immunoglobuline sont identiques dans
leurs parties variables et reconnaissent
donc le même antigène.
2. Rôle des anticorps : la formation des
complexes immuns
Voir activité 24 et doc. 1 p. 316. Les
molécules diffusent de façon radiale dans
Chapitre 11 – L'immunité adaptative 3 / 8

la gélose à partir de chacun des puits. La vitesse de diffusion dépend de la taille des molécules.
L’apparition d’un précipité blanchâtre marque l’agglutination des protéines antigéniques.
Si on avait fait la manipulation avec, dans le puits 1, du sérum provenant d'un lapin non immunisé, on
n'aurait eu aucun arc de précipitation : les anticorps sont produits seulement après un premier
contact avec un antigène (l'albumine de bœuf ici). De plus, ces anticorps sont spécifiques de
l'antigène en question : ils ne forment de complexe immun qu'avec cet antigène.
Les immunoglobulines (anticorps) circulent dans le milieu extracellulaire (milieu intérieur de
l'organisme : plasma sanguin, lymphe), et agissent en se liant spécifiquement aux antigènes qui
ont déclenché leur formation. Antigènes et anticorps fixés dessus constituent le complexe immun.
(Voir aussi doc. 2 p. 317)
3. Elimination des complexes
immuns
Les cellules phagocytaires
(macrophages, granulocytes...),
expriment des récepteurs de la
partie constante des
anticorps, qui leur permettent
de fixer les complexes
immuns. Ils les éliminent alors
par phagocytose (voir doc. 3 p.
317 et ci-contre).
4. Production des anticorps
Voir docs p. 2 de ce polycopié.
Les anticorps sont produits
par des lymphocytes B
sécréteurs = plasmocytes.
De très nombreux clones de
lymphocytes B préexistent avant tout contact avec l'antigène. Chacun porte à sa surface des
immunoglobulines membranaires (ou anticorps membranaires) capables de reconnaître un antigène
et un seul : chaque clone de lymphocytes B est spécifique d'un antigène.
Chapitre 11 – L'immunité adaptative 4 / 8

La reconnaissance d’un antigène donné par un lymphocyte B porteur d’une immunoglobuline
spécifique de cet antigène (« sélection clonale ») entraîne la multiplication de ce lymphocyte
(prolifération) et la formation d’un clone de lymphocytes B ayant la même spécificité.
Les lymphocytes B obtenus se différencient pour certains en plasmocytes sécréteurs d'anticorps et
pour d'autres en lymphocytes B mémoire.
Schéma à connaître : schéma-bilan de la production d'immunoglobulines
Les immunoglobulines sont centrales dans la lutte contre les pathogènes extra-cellulaires.
En ce qui concerne une infection par un virus (ex. virus de la grippe), les immunoglobulines dirigées
contre les protéines virales peuvent bloquer la pénétration des virus dans les cellules, mais ne
peuvent pas agir sur les cellules déjà infectées. Comment celles-ci sont-elles éliminées ? (voir V)
V. Les lymphocytes T cytotoxiques, agents du maintien de l’intégrité des populations
cellulaires
Les cellules infectées par un virus ou un autre agent pathogène intracellulaire exposent à leur surface
des fragments peptidiques issus des protéines du pathogène ; ces fragments sont associés à des
molécules du CMH. Les lymphocytes T cytotoxiques LTc, par leurs récepteurs T spécifiques,
reconnaissent ainsi les cellules infectées. Cette reconnaissance déclenche un mécanisme
d’élimination de la cellule infectée (« baiser de la mort », voir doc. 1 p. 318) : le LTc libère
notamment une protéine appelée perforine, qui s'insère dans la membrane de la cellule à éliminer et
forme des canaux permettant l'entrée et sortie libre d'ions ; cela entraîne la mort de la cellule ; les
débris cellulaires sont ensuite éliminés par les cellules phagocytaires).
Les lymphocytes T cytotoxiques (LTc) portent à leur surface la molécule membranaire CD8. La
production de lymphocytes T cytotoxiques à partir de lymphocytes T CD8 naïfs repose sur des étapes
(sélection, prolifération, différenciation) voisines de celles conduisant à la production de plasmocytes
ou de lymphocytes T auxiliaires (voir doc. 2 p. 319). Comme pour les autres types de lymphocytes,
certaines cellules du clone se différencient immédiatement en LTc, mais d'autres deviennent des
cellules mémoires.
VI. Le S.I.D.A., un effondrement des défenses immunitaires causé par un virus
S.I.D.A. : Syndrome d'ImmunoDéficience Acquise (=A.I.D.S. en anglais)
V.I.H. : Virus de l'Immunodéficience Humaine (=H.I.V.)
Le VIH est un virus fragile qui ne survit pas en dehors de l'organisme. La contamination par le VIH se fait donc
par l'intermédiaire de liquides organiques provenant d'une personne infectée : le sang, le sperme, les
sécrétions vaginales, le lait. Plus de 33 millions de personnes vivent actuellement dans le monde avec une
infection à VIH.
Le VIH est transmis par voie sexuelle, par voie sanguine, ou de la mère à l'enfant (au cours des
derniers jours de la grossesse, au cours de l'accouchement, ou au début de l'allaitement).
Les cellules cibles du VIH sont essentiellement les lymphocytes T CD4. Une fois que le virus a
pénétré dans un de ces lymphocytes, son ARN est rétrotranscrit en ADN grâce à une enzyme du virus
(transcriptase inverse ou rétrotranscriptase). Cet ADN viral s'intègre alors dans l'ADN de la cellule-
hôte. Immédiatement ou après un délai qui peut aller jusqu'à plusieurs années, le génome du V.I.H.
va alors s'exprimer, par transcription et traduction de ses gènes, et provoquer la production de
nombreuses particules virales par la cellule infectée. Ces nouveaux virus bourgeonnent à la
surface de la cellule (qui finit par mourir) et vont infecter de nouvelles cellules cibles. (doc. 1 p. 322)
Ainsi, le V.I.H. entraîne la disparition progressive des lymphocytes T CD4, ce qui empêche la
production d’anticorps contre des agents microbiens variés et l'activation des cellules phagocytaires.
Cela permet l’apparition de maladies opportunistes (infections et cancers). Les conséquences de
l’effondrement des défenses immunitaires prouvent qu’en permanence les mécanismes immunitaires
sont à l’œuvre et montrent le rôle essentiel des lymphocytes T CD4 dans la majorité de ces
réactions.
Chapitre 11 – L'immunité adaptative 5 / 8
 6
7
8
6
7
8
1
/
8
100%