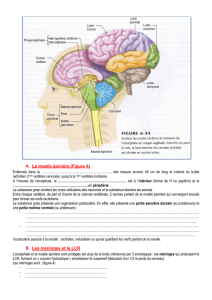
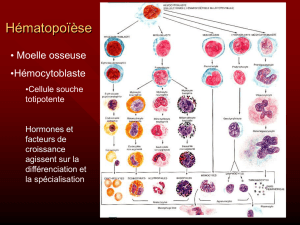
Potentialites hematopoietiques de tissus embryonnaires et adultes

/. Embryo!, exp. Morph.
Vol.
33, 2, pp. 259-278, 1975 ' 259
Printed
in Great
Britain
Potentialites hematopoietiques de tissus
embryonnaires et adultes analysees par greffe a des
poussins irradies
Par JACQUES SAMARUT ET VICTOR NIGON1
Departement de Biologie generate et
appliquee,
Villeurbanne
SUMMARY
Differentiation of
haemopoietic
tissues from embryos and adults
injected into irradiated chickens
Irradiated chicken are injected with haemopoietic tissues from adult or 11-day-old embryos.
Development of stem cells gives rise to well-defined erythrocytic colonies on the surface of
the tibial marrow. Erythropoietic production appears to be similar from adult marrow and
embryonic blood stem cells; production from injected vitelline stem cells seems to be between
3 and 4 times higher than that of adult marrow stem cells. Results are discussed on the basis
of two hypotheses:
- existence of an extramedullary erythropoietic site in the host after vitelline cells grafting;
- development of vitelline stem cells in the host marrow with kinetic patterns different
from those of grafted adult marrow or embryonic blood stem cells.
Anyway, the 11-day-old embryo appears to contain at least two types of blood stem cells
with distinctive properties. Developmental origin, relationship and future of these different
stem cells remain to be analysed.
INTRODUCTION
Au cours du developpement embryonnaire des Vertebres, on observe une
evolution de l'activite erythropoietique caracterisee tant par la sequence des
hemoglobines produites que par la succession des organes erythropoietiques
mis en jeu. Chez le jeune embryon de poulet, la paroi du sac vitellin represente
le principal site erythropoietique. Vers la fin du developpement embryonnaire,
l'activite se concentre, au contraire, dans la moelle, ou elle se poursuit apres
l'eclosion.
Quelles sont les relations genetiques entre ces diverses lignees erythropoi-
etiques qui se succedent au cours du developpement? Selon Moore & Owen
(1967),
les cellules souches hematopoietiques du sac vitellin de l'embryon de
poulet colonisent tous les autres centres hematopoietiques de l'embryon. En
effet, Pinjection de cellules du sang ou du sac vitellin d'un embryon a un
1 Adresse cfauteurs: Departement de Biologie generate et appliquee, 43, Boulevard du
11 novembre 1918 69621 Villeurbanne, France.
i7 EMB 33

260 J. SAMARUT ET V. NIGON
embryon irradie de sexe different et possedant done des structures chromo-
somiques reconnaissables, permet d'identifier une colonisation de la rate, de la
moelle, de la bourse de Fabricius et du thymus par des cellules issues du greffon.
De meme, l'echange de sang etabli par parabiose entre deux embryons de sexes
differents montre une colonisation des centres hematopoietiques de chaque
embryon par les cellules provenant de l'autre embryon et transitant dans la
circulation sanguine (Moore & Owen, 1965).
Chez l'embryon de souris, les travaux de Moore & Metcalf (1970) et Metcalf &
Moore (1971) ont conduit ces auteurs a proposer un modele similaire a celui
propose pour l'embryon de poulet. Les observations de Rifkind, Chui & Epler
(1969) tendent cependant a montrer que rerythropoiese hepatique prend nais-
sance a partir de cellules mesenchymateuses en place dans l'organe lui-meme.
Chez les Batraciens, les greffes d'ilots sanguins triploides sur des embryons
diploides ont mis en evidence l'independance genetique entre les lignees erythro-
poietiques de l'embryon et du tetard (Hollyfield, 1966; Deparis, 1968, 1974).
Au regard de ses contradictions vis a vis des conclusions adoptees par les
autres auteurs, la these de Moore & Owen ramenant l'origine de l'ensemble de
l'erythropoiese du poulet aux cellules du sac vitellin n'apparait pas entierement
convaincante. En effet, s'il existait chez l'embryon un organe different du sac
vitellin et susceptible de produire des cellules souches hematopoietiques, celles-
ci se retrouveraient sans doute egalement dans le sang circulant et partant dans
le sac vitellin. En outre, la duree des experiences faites par ces auteurs est trop
limitee pour permettre de caracteriser pleinement les aptitudes des cellules
greffees. Nous avons done repris des experiences similaires en utilisant comme
receveurs des poussins irradies et en comparant l'effet, sur ces poussins, d'injec-
tions provenant d'organes hematopoietiques de l'embryon et de l'adulte. Chez
un tel hote, la definition des parametres de l'erythropoiese peut, en effet, etre
poussee beaucoup plus loin que chez des embryons.
MATERIEL ET METHODES
Tous les animaux utilises sont de race Leghorn.
Les irradiations sont pratiquees par groupe de quatre poussins maintenus en
position accroupie dans une boite de contention en plexiglass. Ces poussins,
ages de trois semaines, recoivent 760 R le premier jour et 990 R 24 h plus tard.
Le generateur de rayons X est du type Stabilipan Siemens. Le rayonnement est
mesure a l'aide d'un dosimetre Radocon-Victoreen en placant la sonde dans
l'air au centre du volume fictif normalement occupe par un animal. Le debit
de 76 R par minute est obtenu dans les conditions suivantes: distance source-
cible 56,5 cm; filtre 0,2 mm Cu; 170 kV, 20 mA.
La preparation des cellules est effectuee sterilement. Les instruments, la
verrerie et les liquides servant a la preparation des milieux sont sterilises par
chauffage.

Tissus hematopo'ietiques injectes a des poussins irradies 261
Moelle. La moelle est obtenue a partir d'animaux considered comme adultes
et ages de deux mois et demi. Les deux tibias et les deux femurs sont preleves,
puis fendus dans le sens de la longueur. La cavite medullaire est curetee avec
des pinces a bouts recourbes et la moelle est recueillie dans du milieu TC 199 en
solution de Earle (BD Merieux) pH 7,2 tamponne a l'HEPES (Acide 7V-2-
Hydroxyethyl Piperazine-JV-2-Ethanesulfonique, Eurobio) additionne de peni-
cilline (Specilline G 10000 iu/lOOml) et de streptomycine (0,01 g/lOOml). La
moelle est dissociee d'abord a l'aide de pinces fines, puis par passages repetes
dans une seringue de 10 ml sans aiguille. La suspension est filtree sur trois
couches de gaze puis centrifugee 10 min a 300 g a 4 °C. Le culot de cellules est
resuspendu et centrifuge a nouveau dans les memes conditions. La suspension
est enfin denombree au moyen d'une cellule de Malassez et la concentration des
cellules est ajustee, de telle sorte que le volume de suspension injecte ne depasse
pas 0,6 ml. La vitalite des cellules est verifiee par le test du bleu trypan: les
preparations comportent au moins 90 % de cellules vivantes.
Sac vitellin. Les oeufs sont ouverts au lleme jour d'incubation. Avant son
ouverture, la coquille est nettoyee avec un tampon de coton imbibe d'alcool.
L'embryon est recueilli, avec ses annexes, dans du serum physiologique sterile.
La paroi vitelline est decoupee en totalite puis placee dans du tampon phosphate
de Dulbecco (PBS) sans Ca2+ ni Mg2+ pH 7,2 additionne de penicilline (Specil-
line G 10000 iu/100 ml) et de streptomycine (0,01 g/100 ml). Elle est dilaceree a
l'aide de pinces fines. Le milieu est additionne de trypsine (BD-Merieux) a la
concentration finale de 0,25 %. L'ensemble, soumis a une agitation constante par
un barreau magnetique, est incube 30 min dans un bain-marie a 38 °C. En fin
d'incubation, la suspension est homogeneisee par passages repetes dans une
seringue de 10 ml sans aiguille, puis filtree sur trois couches de gaze. Elle est
ensuite lavee trois fois par centrifugations de 15 min a 300 g a 4 °C, dans une
solution PBS. La numeration et la dilution sont effectuees comme pour la
moelle. La preparation contient 90 % de cellules vivantes.
Sang d'embryon. Le sang d'embryon de 11 jours est obtenu par aspiration a
partir de veines vitellines sectionnees. Les cellules sont recuperees dans une
solution PBS sans Ca2+ ni Mg2+ puis centrifugees 10 min a 300 g, a 4 °C. Le
culot est lave une fois et la suspension est denombree; la concentration en
cellules est ajustee comme pour la moelle.
Greffe.
La greffe est effectuee par injection dans une veine de l'aile. Les temoins
recoivent un volume de solution PBS identique au volume de suspension injecte.
Observation des parametres
hematologiqu.es.
Les taux d'erythroblastes poly-
chromatophiles et les myelogrammes sont determines sur 1000 cellules environ
a partir de frottis colores au May Griinwald Giemsa. Les numerations sont
effectuees a l'aide d'une cellule de Malassez. Les animaux sont sacrifies par
decapitation et la moelle d'un tibia est recueillie dans du liquide de Hanks ou
elle est dilaceree puis denombree. L'autre tibia est utilise pour la confection
de frottis.
17-2

262 J. SAMARUT ET V. NIGON
Pour la mesure du volume sanguin, les animaux recoivent une injection
intraveineuse de 0,3 ml d'une solution de Bleu Evans a 50 mg/100 ml. Au bout
de 5 min, le sang est recueilli apres decapitation et la densite optique du serum
est lue a 600 nm.
Comptage des colonies medullaires
erythropo'ietiques
chez les animaux irradies
et greffes. Les animaux sont sacrifies par decapitation. La paroi des tibias est
fendue dans le sens de la longueur au moyen d'une petite scie circulaire de
dentiste. Les os sont: ensuite maintenus dans du liquide de Hanks ou la cavite
medullaire est ouverte et la moelle recueillie integralement avec precaution.
Les colonies erythrocytaires apparaissent a la surface de la moelle sous la
forme de petites taches rouges de 0,5 mm de diametre. Le denombrement des
colonies est rendu plus aise apres fixation de la moelle et coloration des colonies.
Pour cela, le boudin de moelle est plonge dans du liquide de Carnoy pendant
une nuit a 4 °C. Le fixateur est ensuite elimine par deux lavages de 10 min dans
Pethanol a 95°. Les boudins de moelle sont plonges pendant 4 min dans une
solution de benzidine (1 g de benzidine dans 100 ml de methanol), puis dans
une solution d'eau oxygenee 110 volumes diluee 100 fois dans l'ethanol 70°,
jusqu'a l'apparition des colonies colorees en vert a la surface de la moelle.
Leur denombrement est effectue immediatement sous la loupe au grossisse-
mentx 12,5.
Mesure de rendement de la greffe. La technique utilisee est celle decrite par
Siminovitch, McCulloch & Till (1963) pour les CFU spleniques chez la souris.
Des poussins de trois semaines irradies recoivent une injection de 2 x 108
cellules de moelle adulte ou de 5 x 108 cellules de sac vitellin d'embryon de
11
jours. Deux heures apres l'injection, les animaux sont sacrifies et leurs tibias
sont immediatement preleves. La totalite de la moelle d'un tibia est injectee a un
poussin de 3 semaines irradie. Le nombre de colonies par tibia, chez les
receveurs secondaires, est determine 6 jours apres la greffe. Les nombres de
colonies observes sont corriges en tenant compte du fait que 0,05 ml de suspen-
sion reste dans le cone de l'aiguille lors de l'injection.
Le rendement de la greffe est calcule selon les modalites que Ton trouvera
page 270.
RESULTATS
I. Nature des
cellules
grejfees. Caracteristiques des temoins
A. Contamination de
la
preparation de sac
vitellin
par le sang circulant
La preparation effectuee a partir du sac vitellin contient des cellules de la
paroi vitelline proprement dites et des cellules apportees par le sang. Chez
l'embryon de 11 jours, le sang circulant comporte essentiellement des normo-
cytes ainsi que des normoblastes et une faible proportion de megalocytes. On
considere que tous les erythrocytes murs appartiennent a la fraction du sang
circulant. Soient a le pourcentage de ces cellules dans le sang et b leur pour-
centage dans la preparation de sac vitellin. La contamination de celle-ci par les

Tissus hematopo'ietiques injectes a des poussins irradies 263
Tableau 1. Contamination des cellules de sac vitellin
par des cellules du sang embryonnaire
Normocytes
Megalocytes
Normocytes
Megalocytes
Sang
circulant
d'embryon
de 11
jours
(valeurs
moyennes
obtenues
a partir
de trois
embryons)
74,0
6,6
11,2
Sac vitellin
A
Preparation I
A
Cellules
dans la
preparation
28,9
2,1
13
Degre de
contamina-
tion par les
cellules
issues
du sang
39,0
32,0
,8
Preparation II
Degre de
contamina-
tion par les
Cellules cellules
dans la issues
preparation du sang
23,5 33,0
1,9 30,0
12,4
Tableau 2. Composition cellulaire de la moelle chez les animaux
te'moins
et chez les animaux greffes
Cellules erythrocytaires
Erythrocytes murs
Erythroblastes poly-
chromatophiles
Proerythroblastes et
erythroblastes basophiles
Cellules granulocytaires
Cellules lymphocytaires
et monocytaires
Cellules thrombo-
cytaires
Nombre total de cellules
Animaux
A
Non
irradies
25,0
x
10'
13,1x10'
10,2x10'
1,7x10'
5,0x10'
0,4x10'
0,2
x
10'
30,6 x
10'
temoins
Irradies
non
greffes
4,4 xlO7
4,2x10'
0,2x10'
0
0
0,1 x 10'
0
4,5 x 10'
Animaux
Sac
vitellin
8 x 108 cell
9,1 x 107
3,0x10'
5,5x10'
0,6x10'
0,5
x
10'
0,2 x 10'
0,1 x 10'
9,9
x
10'
irradies et greffes avec
Sang Moelle de
d'embryon
2,5 mois
8xl08cell 8xl08cell
— 24,0x10'
— 9,2x10'
— 12,6x10'
— 2,2
x
10'
— 0,7 x 10'
—
1,2x10'
— 0,2 x 10'
2,2x10' 26,1x10'
Tous les animaux sont ages de
3
semaines apres
l'eclosion.
Les animaux irradies (greffes ou
non) sont etudies 6 jours apres
l'irradiation.
Le
nombre
d'animaux
etudies est
de 5
pour chaque
modalite de traitement. Les chiffres correspondent a la population cellulaire par tibia.
Les valeurs presentees pour les animaux greffes sont celles de la population cellulaire issue
du greffon, calculees en deduisant des chiffres totaux les nombres de cellules qui subsistent
dans la moelle des animaux irradies non greffes.
Les diverses valeurs du tableau ont ete calculees en multipliant la frequence des divers types
cellulaires, determined
d'apres
les myelogrammes, par le nombre total de cellules recuperees
par tibia. Ce mode de calcul ne tient pas compte de la proportion de cellules detruites au
cours de la dilaceration de la moelle. On peut estimer que ce taux de destruction est identique
pour toutes les moelles etudiees.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%