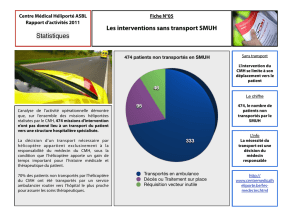
I) Introduction II) CMH : Génétique et structure moléculaire A) Mise

2016-2017 complexe majeur d'histocompatibilité et présentation d'antigène
CMH et présentation d'Antigène
– UE 7 : Immunologie –
Distribution de poly (schémas reportés dans le cours)
Semaine : n°2 (du 06/02/2017 au
10/02/1017)
Date : 06/02/2017
Heure : de 14h00 à
16h00 Professeur : Pr. Bertin
Binôme : n°11 Correcteur : 48
Remarques du professeur
- forum industrie et internat : 8 et 9 mars 2017
- benjamin.bertin-2@univ-lille2.fr
PLAN DU COURS
1/14
I) Introduction
II) CMH : Génétique et structure moléculaire
A) Mise en évidence et définition
B) Structure
C) Diversité génétique
D) Variabilité et déséquilibre de liaison
III) La présentation d' antigènes
A) Lymphocyte : Acteur majeur de la réponse adaptative
B) Fixation de l'Antigène

2016-2017 complexe majeur d'histocompatibilité et présentation d'antigène
I) Introduction:
Immunité adaptative :
Elle fait suite à la réponse immunitaire innée
C'est la reconnaissance et l'élimination sélective des microorganismes spécifiques ou des molécules antigéniques.
Elle se met en place chez les vertébrés. Elle présente 4 caractéristiques majeures :
–spécificité d'antigène : comment s’effectue la présentation d'Ag à l'origine de cette spécificité ?
–énorme diversité
–mémoire immunitaire (vaccination)
–distinction du soi et du non-soi
Elle nous identifie immunologiquement.
Comment interviennent les molécules du CMH dans les greffes ?
Greffe syngénique : souches de souris génétiquement identiques (jumeaux qui partagent le même patrimoine
génétique) → la greffe est acceptée
Allogreffe : greffe chez des individus génétiquement différents au sein d'une même espèce → la greffe est
rejetée
On s'est rendu compte que la base génétique de cette compatibilité tissulaire implique les gènes du CMH, ce qu'on
appelle chez l'homme le système HLA.
Objectifs du cours
:
–Savoir définir ce qu'est le CMH
–Connaitre et comprendre la structure moléculaire des molécules du CMH
–Connaitre et comprendre la diversité génétique des molécules du CMH
–Connaitre et comprendre la fonction des molécules du CMH : la présentation d'antigènes protéiques
Pourquoi ?
Appréhender le CMH dans ses applications cliniques : greffes d'organes et de cellules souches hématopoïétiques
Appréhender le rôle du CMH dans le développement de pathologies (maladies auto-immunes, cancers)
2/14

2016-2017 complexe majeur d'histocompatibilité et présentation d'antigène
II) Le CMH : Génétique et structure moléculaire
A) Mise en évidence et définition
Complexe Majeur d'Histocompatibilité = système HLA (chez l'homme)
COMPLEXE : région d'ADN génomique qui contient plus d'une centaine de gènes codant pour des produits divers
MAJEUR : ses produits sont à l'origine de différences allogéniques importantes entre individus de la même
espèce
Ce qui nous diffère ce sont les groupes sanguins, on parle d'alloantigènes du groupe sanguin et la deuxième grande
diversité sont les gènes du CMH.
HISTOCOMPATIBILITE : ce sont ces régions qui dictent les règles de compatibilité tissulaires lors des greffes (il
faut être histocompatibles pour les groupes HLA)
Le CMH est le marqueur moléculaire de l'identité cellulaire : c'est ce qui nous définit immunogénétiquement
2 rôles
:
–Rôle physiologique : présentation d'antigènes protéiques
–Rôle pathologique : responsable du rejet de greffe tissulaire
Les gènes du CMH se trouvent sur le bras court du chromosome 6.
Cette région génétique est sous-divisée en 3 classes : on parle de CMH de classe I, II et III. Dans ces régions, il y a
des gènes qui n'ont rien à voir avec la réponse immunitaire. On a des gènes majeurs et des gènes mineurs.
Dans la région du CMH classe III on trouve les gènes du complément, des cytokines. Nous n'en reparlerons pas
dans ce cours.
3/14

2016-2017 complexe majeur d'histocompatibilité et présentation d'antigène
B) Structure
CMH I
Le CMH de classe I est une glycoprotéine transmembranaire appartenant à la famille des immunoglobulines.
Attention ce n'est pas pour autant un Ac ! Cela signifie juste que quand on regarde la séquence en AA de cette
chaîne protéique et qu'on la compare à une séquence en AA d'un Ac on se rend compte qu'il y a des similitudes.
Il se trouve à la surface de quasiment toutes les cellules nuclées. (Exceptions : GR, certaines cellules du système
nerveux central, quelques cellules épithéliales très particulières)
C'est un hétérodimère non covalent d'une chaine alpha et de beta 2 microglobuline.Ce sont des interactions
hydrophobes.
Au niveau de la chaine alpha il y a 3 sous-domaines : alpha 1, 2 et 3
Alpha 1 et 2 sont éloignés de la membrane alors que le domaine alpha 3 est proximal.
Le CMH de classe II est également une glycoprotéine transmembranaire appartenant à la famille des
immunoglobulines.
C'est un hétérodimère non covalent d'une chaine alpha avec une chaine beta. Les chaines sont associées entre
elles, mais il n'y a pas de liaison covalente, elles ne sont pas liées physiquement entre elles.
Il est exprimé uniquement à la surface des cellules présentatrices d'antigène
C) Diversité génétique
→ Le CMH est polygénique : pour une même classe de CMH, il existe plusieurs gènes de structure et de
fonctions semblables mais pas identiques.
4/14

2016-2017 complexe majeur d'histocompatibilité et présentation d'antigène
1) CMH I
Il existe 3 gènes majeurs, chacun codant une chaine alpha :
–HLA-A
–HLA-B
–HLA-C
Toutes nos cellules nuclées expriment une version de HLA-A, HLA-B et HLA-C à leur surface.
2) CMH II
Il existe 3 régions (locus) majeures :
–HLA-DP
–HLA-DQ
–HLA-DR
Pour chaque région, on a deux gènes A et B qui codent respectivement pour les chaînes alpha et béta :
–HLA-DPA et HLA-DPB
–HLA-DQA et HLA-DQB
–HLA-DRA et HLA-DRB
Les 6 gènes sont exprimés par nos cellules présentatrices d'antigène.
→ Le CMH est polymorphique : pour chacun des gènes, il existe de nombreux variants alléliques qui vont
différer par leur composition en AA.
Chaque individu reçoit un jeu d’allèle maternel et un jeu d'allèle paternel (au moment de la méiose).
Chaque jeu sera exprimé au sein d'une même cellule : expression codominante.
1) CMH I
On découvre tous les jours de nouvelles allèles, ce sont des chiffres qui changent :
Nombre d’allèles connus en février 2017 : (à ne pas connaitre)
HLA-A : 3830
HLA-B : 4647
HLA-C : 3382
Maman : HLA-A1, HLA-B1, HLA-C1
Papa : HLA-A2, HLA-B2, HLA-C2
= carte d'identité génétique pour le CMH de classe I
A tout instant, chaque cellule exprime les 6 gènes.
5/14
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%