Origine et Evolution de la lignée humaine Biol 301 Myriam HARRY

1
Origine et
Origine et Evolution
Evolution de la lign
de la ligné
ée humaine
e humaine
Biol 301
Myriam HARRY, Université Paris-Sud 11
ET L’HOMME dans tout ça ?
L’homme ne descend pas du singe
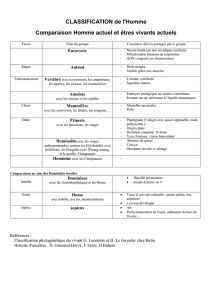
Super-famille
Famille
Sous-famille
Genre
Tribu
Homininae
Homininae Ponginae
Ponginae
Pongo
Pongo
Orang-outang
(1 espèce)
Pongo pygmaeus
Gorilla
Gorilla
Gorille
(1 espèce)
Gorilla gorilla
Hominini
Hominini Gorillini
Gorillini
Hominidae
Hominidae Hylobatidae
Hylobatidae
Hylobates
Hylobates
Gibbon
(12 espèces)
HOMINOÏDES
(ou grands singes)
HOMINOÏDES
(ou grands singes)
Homo
Homo Pan
Pan
Chimpanzé
(2 espèces)
Homme
(1 espèce)
Homo sapiens Pan paniscus
Pan troglodytes
C’est un singe !
Les Primates
Les Primates
• Pouce et gros orteil (sauf humains pour le gros orteil)
opposables aux autres doigts
• Ongles plats à la place des griffes (sauf quelques
petites espèces d’Amérique du Sud)
• Orbite des yeux entièrement cerclé d’os. Yeux
rapprochés sur le devant de la face (vision
binoculaire).
•Museau réduit.
• Cerveau développé.
• Arboricoles en général, parfois terrestres. Tendance
à se redresser.
• La plupart adaptés aux forêts tropicales humides.
51 genres, 168 espèces

2
Singes du Nouveau monde (Platyrhiniens)
Singes de l’Ancien Monde (Catayrhiniens)
Données paléontologiques => 7MA (8-9 MA)
Datation génétique de la divergence chimpanzés/ homme => 5 à 6 MA
Origine des Primates ~ 60 MA
• Petits mammifères insectivores arboricoles (les
plésiadapiformes
)
• fin du Crétacé (-70 MA).
• ressemblaient aux lémuriens actuels
• probablement nocturnes.
• Premiers Simiens (vrais singes) : vers 40 MA.
• Proviendraient de Prosimiens asiatiques (???), peut-être africains.
• Fossile le plus ancien connu (Siamopithecus, 40 MA) a été
découvert en Thaïlande,
• Les simiens ont surtout prospéré en Afrique.
.Séparation des singes du Nouveau monde (Platyrhiniens)
de ceux de l’Ancien Monde (Catayrhiniens) : vers 40 MA.
Beaucoup d’espèces disparaissent suite à l’important
refroidissement du début de l’oligocène.
Les premiers Hominoïdes (grands singes)
Les premiers Hominoïdes (grands singes)
La séparation entre Cercopithécoïdes (singes avec queue) et
Hominoïdes (grands singes) se produit en Afrique il y a environ
20-25 MA.
Plus ancien des grands singes connu = Proconsul
Pas de queue, mais locomotion semblable à celle des
cercopithèques.
À partir de 20 MA : le climat devient plus sec en Afrique de l’Est;
la savane boisée gagne sur la forêt tropicale humide.
Ces conditions profitent aux grands singes (Hominoïdes) aux
dépens des petits singes (Cercopithécoïdes).
Les grands singes constituent même, un moment, 90% des
espèces de singes (c’est l’inverse aujourd’hui).
L’âge d’or des grands singes
L’âge d’or des grands singes

3
Premiers pas vers l’humanité
Premiers pas vers l’humanité
Trois principales différences entre les grands singes et
l’humain
•Volume du cerveau
• Mode de locomotion
• Langage
• Utilisation d’outils
(certains grands singes
utilisent des outils
simples)
Volume du cerveau
Volume du cerveau
Homme : volume important (~1400 cc)
et face réduite.
Grands singes : volume environ quatre
fois plus faible (350 à 400 cc).
Homme : réduction de la mâchoire
=> formation du menton
Homme Chimpanzé
Bébé humain
Bébé
chimpanzé
Implication de gènes du
développement
=> Conservation chez l’humain
à l’état adulte des caractères
physiques typiques de
l’enfance (néoténie)
Adultes
Ressemblance du crâne d’un
humain adulte à celui d’un bébé
chimpanzé.
Mode de locomotion
Mode de locomotion
Grands singes :
- parfois bipèdes (environ 10% à 20% du
temps chez les chimpanzés), mais peu
adaptés à ce mode de locomotion.
- se déplacent surtout à quatre pattes ou
suspendus par les bras aux branches.
Humains : uniquement bipèdes
(sauf les bébés !).
Humains :
Colonne vertébrale
présentant 4 courbures
homme
grand singe
Adaptations du squelette à la
bipédie
Adaptations du squelette à la
bipédie
Colonne vertébrale
L’adaptation à grimper et à se
suspendre aux branches
prédispose à la bipédie.
mêmes muscles des hanches et
du bas du dos sollicités pour
grimper et pour marcher.

4
Adaptations du squelette à
la bipédie
Adaptations du squelette à
la bipédie
Bras et jambes
Humains :
Les bras sont plus
courts que les jambes
La morphologie des bras et
des épaules chez les
humains découle de
l’adaptation de leurs
ancêtres à se suspendre aux
branches.
Humains :
Bassin plus large et
plus robuste (doit
supporter presque
tout le poids du
corps en position
redressée).
Sacrum plus robuste
grand singe homme
Bassin
Fémur
Humains :
- Fémur et col du fémur
plus longs
- Fémur est orienté
différemment par
rapport au bassin
Grands singes :
- Position du trou
occipital plus reculée
grand singe
homme
- Canines très
développées (surtout
chez les mâles)
- Dentition en « U »
Position du trou occipital
et dents
Pieds
Chimpanzé
Homme
Humains :
-gros orteil dans le
prolongement du pied
- pied non préhensile (le gros
orteil ne peut pas s’opposer aux
autres)
- Pied présentant une voûte
plantaire
gros orteil
Evolution de la lign
Evolution de la ligné
ée humaine
e humaine

5
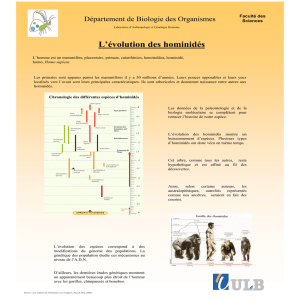
Données paléontologiques
Australopithecus afarensis
= Lucy
Homo neanderthalensis =
Homme de Neandertal
Homo sapiens =
Homme de Cro-Magon
Homo erectus
Homo habilis
Vision linéaire et ancienne
Et fausse
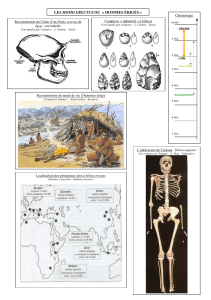
Toumaï
Sahelanthropus tchadensis
Touma
Toumaï
ï
Sahelanthropus
Sahelanthropus tchadensis
tchadensis
-Découvert par Ahounta Djimdoumalbaye
de l’équipe de Michel Brunet
- juillet 2001
- dans le désert du Djourab, au nord du
Tchad (Afrique centrale)
- crâne, quelques dents, fragments de
mâchoire.
- Datation : 7 MA (datation relative et
datation absolue au 10béryllium).
Toumaï signifie « espoir de vie » en langue Goran
Toumaï
Sahelanthropus tchadensis
Touma
Toumaï
ï
Sahelanthropus
Sahelanthropus tchadensis
tchadensis
Taille : 1 à 1,3 m
Trou occipital témoignant
d’une position dressée,
probablement bipède
Autres caractéristiques
typiques des hominidés :
face plus aplatie que celle
des grands singes
Capacité crânienne proche
de celle du chimpanzé
Ancêtre de l’homme ?
ou un ancêtre commun aux grands singes actuels et aux
humains ?
Orrorin tugenensis
Orrorin
Orrorin tugenensis
tugenensis
Découvert en janvier 2001 au Kenya
(Afrique de l’Est) quelques mois avant Toumaï
Fragment de mâchoire, restes du fémur et de
l’humérus, quelques dents, une phalange
Taille : 1,25 et 1,5 m
Datation : 6 MA
Mâchoire primitive comme celle des singes
Fémur indique nettement une locomotion bipède
(longueur, forme, orientation du col)
Humérus et phalange très allongée (comme celle des grands singes)
témoignent une aptitude à la suspension
Australopithecus afarensis
Australopithecus
Australopithecus afarensis
afarensis
Nombreux fossiles retrouvés dont Lucy
(AL 288-1), un squelette complet à 40%
découvert en 1974 par Donald Johanson,
Maurice Taieb et Yves Coppens sur le site
des Afars en Éthiopie
Taille entre 1 et 1,5 m
Datation : 4 et 3 MA
Lucy
Volume crânien ~ celui des
grands singes : 450 cc
Locomotion bipède, mais
moins spécialisée que celle
des humains modernes.
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%