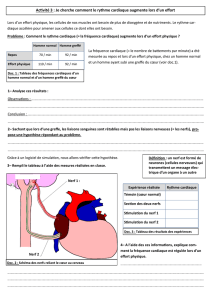
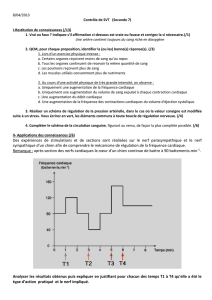
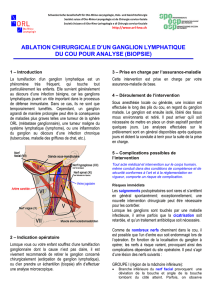
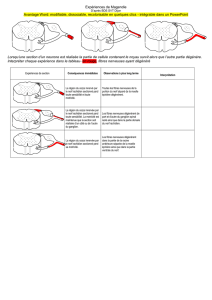
Thesis - Archive ouverte UNIGE

Thesis
Reference
Recherches sur le développement des nerfs craniens chez les
lézards
BÉRANECK, Edmond
BÉRANECK, Edmond. Recherches sur le développement des nerfs craniens chez les
lézards. Thèse de doctorat : Univ. Genève, 1884
DOI : 10.13097/archive-ouverte/unige:21776
Available at:
http://archive-ouverte.unige.ch/unige:21776
Disclaimer: layout of this document may differ from the published version.
1 / 1

RECHERCHES
SUR
LE
DÉVELOPPEMENT
DES
NERFS
CRANIENS
CHEZ
LES
LÉZARDS
PAR
E.BÉRANECK
GENÈVE
1 M P R 1
MER
1
EJ
CH
ARLES
rs
CHU
CH
A(R
DT
1884

La
Fam{
lté
des
Sciences
autorise l'impression
de
la
présente
thèse)
sans exprimer d'opinion sur
les
proposi-
t-ions
qui y sont renfermées.
Le Doyen
de
la
Faculté :
G.
ÜLTRAMARE.
Genève,
Je
3 juillet 1884.

RECHERCHES
SUR
LE
D~VELOPPEMENT
DES
NERFS
CRANIENS
CHEZ
LES
LÉZARDS
Jusqu'à présent
les
embryologistes
qui
·se
sont
occupés
du
développement
du
système
nerveux
chez
les
vertébrés
ont porté
leurs
recherches
plutôt sur
la
formation
des
nerfs
spinaux
que
sur
celles
des
nerfs
crâniens
et,
à part
les
recherches
classiques
de
BALFOUR
dans
sa
«
Monogra-
phie
sur
le
développement
des
poissons
Elasmobranches,
»
et
de
MARSHALL
dans
son
«
Étude
sut'
le
développement
des
nerfs
crâniens
du
poulet,
»
puis
sur «
Les
cavités
céphaliques
et
les
nerfs
des
Élasmobt'anches,,
»
nous
ne
possédons
que
relativement
peu
de
renseignements sur
la
formation
des
nerfs
crâniens
dans
les
différentes
classes
des
vertébrés.
C'est
ce
qui
m'a
engagé
à
faire
de
cette
question
une
étude
aussi
complète
qu'il
m~a
été
possible.
Mes
recherches
ont
été
faites
dans
le
laboratoire
d'embryogénie
de
l'Uni-
versité
de
Genève,
et
je
tiens
ici
à remercier tout parti-
culièrement
Monsieur
le
professeur
FoL,
qui
non
seule-
ment m'a
dirigé
dans
le
choix
de
mon
sujet
et
dans
la
marche
de
mori
travail,
mais
encore a bien
voulu
me

4
E.
BÉRANECK.
fournir
tous
les
matériaux nécessaires, matériaux
longs
et
difficiles
à
se
procurer.
Ces
recherches ont été
faites
sur
le
lézard ordinaire
(
Lacerta
agilis),
et
si
sur
le
conseil
du
profeRseur
FoL
j'ai
choisi
comme
sujet d'étude
un
représentant
de
la
classe
des
Reptiles, c'est
que
nos
connaissances touchant
le
développement
des
nerfs crâniens dans
cette
classe
des
vertébrés sont pour ainsi dire nulles.
J'ai étudié un certain nombre
de
stades, cependant je
n'ai
pu
me
procurm·
les
stades
les
plus
inférieurs
dn
développement ni
des
embryons correspondant aux der-
nières
phases
de
l'évolution embryonnaire
dn
lézard.
Lr.s
embryons ont été durcis par
le
liquide
de
KLEINENBERG
(acide piero-sulfurique), puis successivement traités par
l'alcool à 70°, 90°
et
absolu;
ils
ont élé
colorés
par
le
· carmin-borax
de
GRENACHER
qui
m'a donnà
de
bons
résultats. J'ai étudié 9 stades
de
développement
du
lézard
dont quatre feront
plus
spécialement l'objet
de
ce
travail,
Ces
4 stades seront
désignés
par
les
numéros 4, 6, 9,
'10,
et sont représentés par
les
figures
1,
2,
3
et
4
de
la
plan-
che
I.
Il
m'est tout à fait impossible
de
donnm·
l'âge
même
approximatif
de
ces
embryons. Un
des
seuls
Cf'itériums
qui
puisse
~ervir
à
la
détermination
de
l'âge relatif
de
ces
embryons par rapport
les
uns
aux.
autres étant
la
lon-
gueur,
je
vais
indiquer
dn
moins pour
les
quatre stades
principaux
les
chiffres
exprimant leur longueur
en
mil-
limètres.
L'embryon
du
stade
no
IV
a..
3mm
6
de
longueur.
L'embryon
du
stade U0
Vl
a..
5mm
9 »
L'embryon
du
stade
n°
IX
a..
9mm
5
>>
L'embryon
du
stade
n°
X a
..
27mm
5 »
La
différence
que
l'on constate entre
le
stade IX
et
le
stade X peut paraître
excesgive,
mais
il
faut tenir compte
que
dans
le
stade
IX
la· queue
est
encore enroulée sur
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
1
/
97
100%