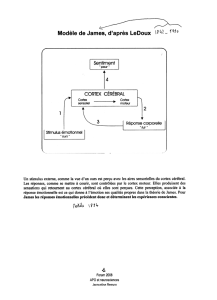
Combiner les apprentissages motivés et associatifs - TEL (thèses

Combiner les apprentissages motiv´es et associatifs
Maxime Carrere
To cite this version:
Maxime Carrere. Combiner les apprentissages motiv´es et associatifs. Autre [cs.OH]. Universit´e
de Bordeaux, 2016. Fran¸cais. <NNT : 2016BORD0191>.<tel-01399886>
HAL Id: tel-01399886
https://tel.archives-ouvertes.fr/tel-01399886
Submitted on 21 Nov 2016
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

THÈSE
PRÉSENTÉE À
L’UNIVERSITÉ DE BORDEAUX
ÉCOLE DOCTORALE DE MATHÉMATIQUES ET
D’INFORMATIQUE
par Maxime Carrere
POUR OBTENIR LE GRADE DE
DOCTEUR
SPÉCIALITÉ : INFORMATIQUE
Combiner les apprentissages motivés et associatifs
Date de soutenance : 11 octobre 2016
Devant la commission d’examen composée de :
Frédéric Alexandre Directeur de recherche, Inria . . . . . . . . . . . . . . . . . . . . . . . . . . . Directeur
Peter Dominey . . . . . . Directeur de recherche, SBRI . . . . . . . . . . . . . . . . . . . . . . . . . . Rapporteur
Mehdi Khamassi . . . . Chargé de recherche HDR, ISIR . . . . . . . . . . . . . . . . . . . . . . . Rapporteur
Pierre-Yves Oudeyer Directeur de recherche, Inria . . . . . . . . . . . . . . . . . . . . . . . . . . . Examinateur
Aline Desmedt . . . . . . Maître de conférence HDR, Neurocentre Magendie . . . . . Examinateur
Julien Vitay . . . . . . . . . Associate Professor, Chemnitz University of Technology Examinateur
2016


Résumé Pour pouvoir être autonomes dans un environnement complexe,
les humains comme les systèmes artificiels doivent posséder un apprentissage
souple et capable de s’adapter au changement. Dans cette thèse, nous nous in-
téressons à comment cette autonomie peut être obtenue par interactions entre
les différents systèmes d’apprentissage de notre cerveau. Pour cela, nous modé-
lisons dans une approche inspirée de la biologie le comportement de certaines
des parties du cerveau impliquées dans les apprentissages répondant et opérant,
et observons comment leurs interactions permettent un apprentissage flexible
dans des tâches impliquant des changements comme l’extinction et le reversal.
Title Combining associative and motivated learning.
Abstract In a complex environment, humans and artificials systems need a
flexible learning system to adapt themselves to situations which can change. In
this thesis, we study how autonomy can be the result of interactions between
the different learning systems of our brain. In particular, in a biologically-
inspired approach, we model different parts of the brain involved in respondant
and operant conditioning, et show how their interactions can promote flexible
learning in tasks in which situation can change, like extinction or reversal.
Keywords Learning, Biologically-inspired, Neuroscience, Amygdala, Cor-
tex, Striatum, Neuromodulation.
Mots-clés Apprentissage, Bio-inspiré, Neuroscience, Amygdale, Cortex, Stria-
tum, Neuromodulation.
Laboratoire d’accueil Laboratoire Bordelais de Recherche en Informatique
(LaBRI). 351, cours de la Libération, Talence, France
Combiner les apprentissages motivés et associatifs iii

iv Maxime Carrere
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
1
/
132
100%