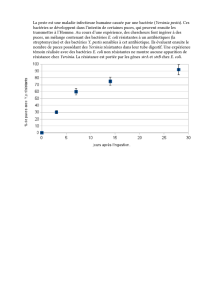
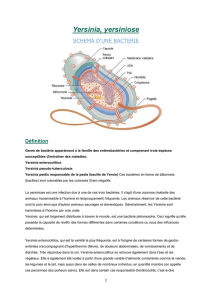
la pseudotuberculose a yersinia pseudotuberculosis en

ECOLE NATIONALE VETERINAIRE D’ALFORT
ANNEE 2004
LA PSEUDOTUBERCULOSE
A YERSINIA PSEUDOTUBERCULOSIS
EN PARCS ZOOLOGIQUES
THESE
Pour le
DOCTORAT VETERINAIRE
Présentée et soutenue publiquement
devant
LA FACULTE DE MEDECINE DE CRETEIL
Le
Par
Laurence, Magali CRENN
Née le 30 avril 1978 à Châtillon-sous-Bagneux (Hauts-de-Seine)
JURY
Président :
Professeur de la Faculté de Médecine de CRETEIL
Membres :
Directeur : M. BOULOUIS
Professeur à l’Ecole Nationale Vétérinaire d’Alfort
Assesseur : M. COURREAU
Professeur à l’Ecole Nationale Vétérinaire d’Alfort


REMERCIEMENTS
A Monsieur le Professeur de la Faculté de Médecine de Créteil
qui nous a fait l’honneur de présider notre jury de thèse
A Monsieur BOULOUIS
Professeur à l’Ecole Nationale Vétérinaire d’Alfort
pour avoir accepté la direction et le sujet de cette thèse
A Monsieur COURREAU
Professeur à l’Ecole Nationale Vétérinaire d’Alfort
pour son aide précieuse tout au long de nos années à l’Ecole
A Monsieur MOISSON
initiateur du sujet de cette thèse
pour son accueil chaleureux et le travail partagé
A tous les vétérinaires ayant répondu aux questionnaires
pour le temps consacré et leur précieuse collaboration
Au personnel du Parc Zoologique et Botanique de Mulhouse
pour les bons moments passés en leur compagnie durant tout mon stage


A mes parents,
sans qui rien de tout cela n’aurait pu être,
merci pour le soutien tout au long du chemin,
c’est à vous que je dois d’être vétérinaire aujourd’hui.
A mes grand-parents,
vous êtes mes racines : grâce à vous, je sais d’où je viens.
A mon grand-père surtout,
qui est parti si brutalement l’année de mon concours .
A Sandrine, ma sœur,
parce que nos vocations respectives ne sont pas si différentes.
A Maeva, ma nièce et filleule,
j’espère arriver à te transmettre l’amour et le respect de la vie animale,
pour que tu n’aies pas à connaître un monde dénaturé par les erreurs de tes aînés.
A Yaël et Louisa,
petites fleurs de demain.
A ma famille du Jura,
pour m’avoir fait si vite une place auprès d’elle.
A Gisèle et Baptiste particulièrement,
parce que, grâce à vous,
nous sommes une famille recomposée vraiment réussie.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
1
/
156
100%