poster montpellier locus du sexe marine VF

Caractérisation du locus du sexe de l’algue
brune modèle Ectocarpus siliculosus
E. siliculosus, un organisme modèle pour
les algues brunes
Facile à cultiver (boîtes de Petri)
Cycle de vie au laboratoire (3 mois pour le cycle
complet)
Possibilité de croisements génétiques
Beaucoup d’écotypes disponibles
Petit génome (200 Mbp - 1000 Mbp pour Fucus)
Bien décrit dans la littérature
Cohabite avec des macroalgues exploitées (Laminaires)
MAIS: pas de différence phénotypique entre les sexes
Les récents progrès dans les techniques de séquençage permettent aujourd’hui d’étudier l’évolution des chromosomes sexuels ou des régions du
déterminisme sexuel chez plusieurs organismes modèles, aussi bien chez les mammifères que chez les plantes dioïques, les poissons et les champignons. Les
systèmes génétiques de déterminisme du sexe ont évolué de manière indépendante selon les taxons mais ils montrent une convergence évolutive: apparition
d’une petite région du déterminisme sexuel sur un chromosome ordinaire, suppression progressive de la recombinaison entre chromosomes homologues, et
éventuellement dégénérescence et réduction de la taille d’un des deux chromosomes sexuels (Charlesworth et al., 2005). Les chromosomes sexuels humains X
et Y (Fig. 1a) ont suivi ces étapes et sont maintenant bien différenciés, mais certains loci « mating type »- comme ceux de Chlamydomonas reinharditi -
présentent des caractères de chromosomes sexuels primitifs (Fig. 1b). Les algues brunes forment un groupe extrêmement intéressant pour l’étude de
l’évolution des chromosomes sexuels car on y trouve une grande diversité de systèmes sexuels, allant de l’isogamie à l’oogamie en passant par différents
degrés de différentiation sexuelle. L’algue brune filamenteuse Ectocarpus siliculosus présente un intérêt particulier car i) elle possède un système sexuel très
simple: les gamètes sont morphologiquement identiques (isogamie) mais la femelle produit une phéromone qui attire le mâle (anisogamie comportementale)
et ii) c’est un organisme haploïde-diploïde (Fig. 2) or, si le déterminisme sexuel est bien étudié chez les espèces diploïdes, il l’est très peu chez les espèces
haploïdes ou haploïdes-diploïdes. La caractérisation du système sexuel d’E.siliculosus pourrait ainsi permettre de mieux comprendre les premières étapes de
l’évolution des systèmes sexuels.
INTRODUCTION
M Robuchon1,2, S Dittami4, AF Peters5, D Roze2, M Valero2, C Destombe2, JM Cock1, JH Bothwell3, Coelho SM1
ETUDE DE LA RECOMBINAISON
Le locus du sexe mâle d’E.siliculosus est une région non recombinante d’au moins 0.8Mbp située sur le groupe de liaison
30. Il occupe approximativement 20% de ce groupe de liaison.
Des études sur le niveau de polymorphisme neutre au sein et en dehors du locus du sexe sont prévues sur des
gamétophytes mâles issus de populations naturelles du Pérou et du Chili afin de voir jusqu'à quel niveau les mécanismes
de sélection ont réduit le polymorphisme au sein du locus du sexe et des régions avoisinantes (Charlesworth et al., 1993).
Une souche femelle est en cours de séquençage, ce qui permettra de comparer les génomes mâle et femelle en
particulier au niveau du groupe de liaison 30.
CONCLUSION ET PERSPECTIVES
IDENTIFICATION DU LOCUS DU SEXE
Des expériences sur puces à ADN (Dittami et al., soumis) ainsi que les
ressources générées par le projet de séquençage du génome d’un
gamétophyte mâle d’E. siliculosus (Cock et al., 2010) ont permis d’identifier
plusieurs zones du génome issues de sept supercontigs présentes
uniquement chez les mâles, notamment la totalité du supercontig 68 (Fig. 3).
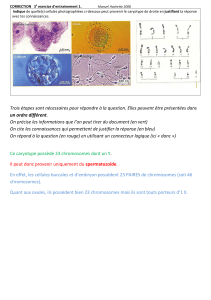
Des tests de liaison au sexe (Fig. 4) ont montré que le supercontig 68 mais
aussi le supercontig 285 étaient mâle-spécifiques et, grâce à la carte
génétique développée pour E.siliculosus (Heensch et al., in press), ces
supercontigs ont été localisés tous les deux sur le groupe de liaison 30 . De
manière intéressante, le groupe de liaison 30 présente un fort taux
d’éléments répétés et de transposons ainsi qu’une faible densité de gènes qui
sont des caractéristiques emblématiques des chromosomes sexuels (Ming
and Moore, 2007).
1Equipe Génétique des algues, UMR 7139, CNRS-UPMC , Station Biologique de Roscoff, France
2Equipe Biologie évolutive et diversité marine, UMR 7144, CNRS-UPMC Station Biologique de Roscoff, France
3Queen's University Belfast , UK
4Equipe Stress abiotique et génomique fonctionnelle des macroalgues, UMR 7139, CNRS-UPMC , Station Biologique de Roscoff, France
5Bezhin Rosko, Roscoff, France
-3
-2
-1
0
1
2
3sctg 69 sctg 68 (red) sctg 178
Log2-ratio
Figure 3: Extrait du signal d’hybridation entre le génome d’une
souche mâle et le génome d’une souche femelle. En rouge, une
région du génome femelle qui ne s’hybride pas avec le génome mâle Figure 4: Le supercontig 68 co-ségrège avec le sexe. F=femelle, M=mâle
Figure 1: Diagramme des différents types de
chromosomes sexuels, d’après Bergero et Charlesworth,
2009.
Lignes épaisses: régions recombinantes, lignes fines:
régions non recombinantes. a) Modèle XY classique dans
lequel le Y est majoritairement constitué d’une région
mâle spécifique dégénérée contenant moins de gènes que
la région homologue sur le X; b) Modèle où le Y contient
une petite région avec les propriétés d’un chromosome
sexuel et est normal ailleurs
Le but de ce projet est d’identifier et de caractériser la région du déterminisme sexuel d’ E. siliculosus
Figure 2: Cycle de vie haploïde-diploïde d’E.siliculosus
Le sexe est exprimé en phase haploïde chez les gamétophytes tandis que le
sporophyte diploïde possède à la fois les régions mâle et femelle
Marqueur du spct 68
Contrôle interne de PCR
M F M F M M
Références
•Bergero R and Charlesworth D (2009). The evolution of restricted
recombination in sex chromosomes. Trends in Ecology & Evolution,24,94-102.
•Charlesworth D, Charlesworth B and Marais G (2005). Steps in the evolution of
heteromorphicsex chromosomes. Heredity, 95(2): 118-128.
•Charlesworth B, Morgan MT, Charlesworth D (1993). The effect of deleterious
mutations on neutral molecular variation. Genetics 134(4):1289-1303.
•Cock JM, Sterck L, Rouze P, Scornet D, Allen AE, Amoutzias G, Anthouard V,
Artiguenave F, Aury JM, Badger JH et al. (2010). The Ectocarpusgenome and the
independent evolution of multicellularity in brown algae. Nature 465(7298):617-
621.
•Dittami SM, Proux C, Rousvoal S, Coelho SM, Peters AF, Cock JM, Coppée JY, Boyen
C, Tonon T. Comparative genome hybridization of different strains and ecotypes of
Ectocarpus reveal high intra-generic variability (submitted to New Phytologist).
•Heesch S, Cho GY, Peters AF, Le Corguillé G, Falentin C, Boutet G, Coëdel S, Jubin
C, Samson G, Corre E, Coelho SM and Cock JM (2010). A sequence-tagged genetic
map for the brown alga Ectocarpus siliculosus provides large-scale assembly of the
genome sequence. New Phytologist (in press).
•Ming R, Yu Q, Moore PH (2007). Sex determination in papaya. Semin Cell Dev
Biol. 18(3):401-8.
L’étude de la recombinaison s’est faite sur 2000 parthénosporophyte issus
de 2000 méioses différentes, à l’aide de trois marqueurs: deux marqueurs
situés chacun à une extrêmité du supercontig 68 et un marqueur du
supercontig 285.
Aucun évènement de recombinaison n’a été détecté, ni entre les deux
extrêmités du supercontig 68, ni entre le supercontig 68 et le supercontig
285. Ceci est cohérent avec le fait qu’il s’agit du locus du sexe et implique que
les supercontigs 68 et 285 sont très proches physiquement, sans doute
accollés, formant ainsi une zone non recombinante continue d’environ
0.8Mpb.
1
/
1
100%