Cours 5

Partie 2
4 et 5.3.09

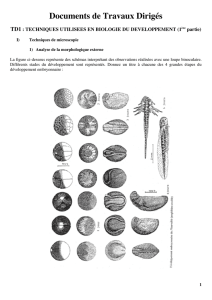
Une expérience ancienne, mais déterminante !
Revenons un peu en arrière….
Sur la capacité inductrice de
la lèvre dorsale du blastopore

Expérience de Spemann
et Mangold (1924)

Analyse histologique
Conclusion:
L’analyse histologique met en évidence deux axes embryonnaires
complet.
La distribution du pigment révèle les contributions respectives de
l’implant et des tissus de l’hôte:
-chorde
-région proximale des somites
-toit de l’archentéron de l’embryon secondaire
La lèvre dorsale est
un tissu inducteur =
centre de Spemann

Molécules inductrices sécrétées
(cellules inductrices)
(cellules cibles)
Récepteurs spécifiques
Signalisation > protéines cytoplasmiques
Réponse cellulaire
Conclusion Les cellules de l’embryon en segmentation
sont déterminées.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
1
/
54
100%