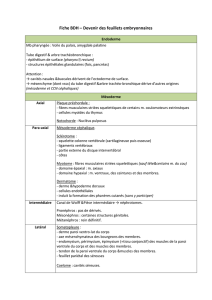
La théorie des feuillets : implications embryologiques - iPubli

HYPOTHÈSE
Il :

n'



1
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%