Apparition et évolution de la résistance bactérienne aux antibiotiques

Apparition et ´evolution de la r´esistance bact´erienne aux
antibiotiques
J.F. Guillot
To cite this version:
J.F. Guillot. Apparition et ´evolution de la r´esistance bact´erienne aux antibiotiques. Annales
de Recherches V´et´erinaires, INRA Editions, 1989, 20 (1), pp.3-16. <hal-00901839>
HAL Id: hal-00901839
https://hal.archives-ouvertes.fr/hal-00901839
Submitted on 1 Jan 1989
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

Article
de
synthèse
Apparition
et
évolution
de
la
résistance
bactérienne
aux
antibiotiques
J.F.
Guillot
INRA,
Station
de
pathologie
aviaire
et
de
parasitologie,
CR
de
Tours-Nouzilly,
37300
Monnaie,
France
(reçu
le
28-4-1988,
accepté
le
7-6-1988)
Résumé ―
L’utilisation
thérapeutique
croissante
d’antibiotiques
a
conduit
à
l’émergence
rapide
de
souches
bactériennes
multirésistantes.
Les
souches
antibiorésistantes
sont
d’abord
apparues
en
milieu
hospitalier;
récemment,
leur
rôle
pathogénique
dans
certaines
infections
chez
l’homme
et
l’animal
et
leur
présence
dans
la
flore
digestive
humaine
ont
été
observés.
De
plus,
des
bactéries
résistantes
ont
été
isolées
de
l’environnement.
antibiotiques -
souches
bactériennes
antibiorésistantes -
bactériologie
Summary ―
Discovery
and
evolution
of
microbial
resistance
to
antibiotics.
Concomitant
with
antibiotic
use
has
been
the
appearance
of
resistance
bacteria
which
seem
able
to
emerge
as
rapid-
ly
as
new
antibiotics
are
introduced.
Although
initially
encountered
in
hospitals,
resistant
bacteria
are
now
being
detected
as
causes
of
human
and
animal
infections
and
as
colonizers
of
the
gut.
Moreover,
resistant
bacteria
are
isolated
from
the
environment.
antibioresistance -
bacteriology -
antibiotics
Introduction
Depuis
leur
découverte,
les
antibiotiques
se
sont
révélés
très
précieux
dans
la
lutte
contre
les
maladies
d’origine
bactérienne
touchant
l’homme
et
les
animaux.
En
éle-
vage,
associés
aux
vaccinations
et
à
une
amélioration
des
conditions
hygiéniques,
ils
ont
fortement
contribué
au
développe-
ment
de
l’élevage
dit
«industriel».
A
cet
égard
les
volailles
sont
intéressantes,
car
l’élevage
avicole
a
été
le
premier
à
s’in-
dustrialiser
et
c’est
aussi
chez
le
poulet
qu’a
été
découverte
une
autre
propriété
des
antibiotiques :
l’effet
promoteur
de
croissance.
Mais
le
monde
bactérien
s’est
adapté
aux
antibiotiques
et
cela
s’est
traduit
par
l’émergence
de
souches
résistantes
chez
l’homme,
chez
les
animaux
et
dans
l’envi-
ronnement.
L’existence
de
ces
bactéries
résistantes
a
des
conséquences
sur
la
thérapeutique,
la
santé
publique
et
l’hygiè-
ne
de
l’environnement.

Evolution
de
la
consommation
des
antibiotiques
Découverte
en
1928
par
Fleming,
con-
centrée
et
purifiée,
par
Chain
et
al.
vers
1940,
la
pénicilline
commença
à
être
notablement
utilisée
en
1943
pour
traiter
les
so,ldats
des
armées
alliées.
Entre
1941
et
1950,
les
premiers
représentants
des
principales
familles
antibiotiques
furent
découverts :
strepto-
mycine,
tyrothricine,
chloramphénicol,
tétracycline.
Ces
nouvelles
molécules
permirent
d’élargir
le
spectre
d’activité
des
antibiotiques
et
d’améliorer
ainsi
la
lutte
contre
les
maladies
bactériennes.
Depuis
1950,
la
liste
des
antibiotiques
n’a
cessé
de
s’allonger.
Entre
1951
et
1959,
période
la
plus
féconde,
40
à
60
molé-
cules
nouvelles
étaient
décrites
chaque
année.
Parallèlement
à
l’usage
humain,
l’utili-
sation
de
ces
substances
en
médecine
vétérinaire
s’est
rapidement
développée.
Dès
1950,
les
traitements
et
la
prévention
avec
les
tétracyclines,
la
streptomycine,
les
sulfamides
et
le
chloramphénicol
ont
été
pratiqués
chez
les
animaux
d’élevage.
En
1946,
Moore
et
al.
découvrent
chez
le
poulet
l’effet
promoteur
de
croissance
de
faibles
doses
de
sulfamides
et
de
streptomycine.
Cet
effet
zootechnique-
ment
favorable
a
été
retrouvé
avec
d’autres
antibiotiques :
tétracycline
et
antibiotiques
polypeptidiques.
Dès
lors,
l’incorporation
d’antibiotiques
à
faible
dose
(de
quelques
ppm
à
quelques
cen-
taines)
dans
l’aliment
destiné
aux
ani-
maux
de
rente
est
devenue
une
pratique
courante.
Ces
différents
usages
des
antibio-
tiques,
thérapeutique
et
prophylactique
chez
l’humain,
thérapeutique,
prophylac-
tique
et
zootechnique
chez
l’animal
et
les
effets
bénéfiques
obtenus
expliquent
faci-
lement
l’augmentation
constante
de
la
consommation
de
ces
substances
(Fig.
1).
).
L’usage
des
antibiotiques
en
médecine
vétérinaire
est
loin
d’être
négligeable
(Tableaux
I et
11).
Environ
40%
des
anti-
biotiques
produits
sont
utilisés
chez
les
animaux.
L’administration
des
antibio-
tiques
dans
l’aliment
correspond
essen-
tiellement
aux
additifs
promoteurs
de
croissance.
Cet
usage
des
antibiotiques
chez
les
animaux
et
plus
précisément
celui
des
additifs
promoteurs
de
croissance
a
pré-
occupé,
dès
les
années
60,
les
médecins
et
l’opinion
publique
dans
la
mesure
où
les
antibiotiques
utilisés
pourraient
favori-
ser
l’apparition
de
bactéries
résistantes
transmissibles
à
l’homme.
Certains
pays
ont,
malgré
l’absence
de
preuves
scienti-
fiques
précises
sur
les
risques,
envisagé
une
réglementation.
En
Grande-Bretagne,
dès
1969,
le
rap-
port
Swann
assez
théorique
classe
les
antibiotiques
promoteurs
de
croissance
en
2
groupes,
ceux
utilisés
en
thérapeu-
tique
et
les
«non-thérapeutiques».
Il
vise

à
restreindre
l’usage
comme
additifs
d’an-
tibiotiques
utilisés
en
thérapeutique
en
préconisant
l’autorisation
de
produits
éco-
nomiquement
valables,
de
peu
d’intérêt
en
thérapeutique
humaine
ou
animale,
ne
gênant
pas
l’efficacité
de
la
thérapeutique
et
ne
sélectionnant
pas
de
bactéries
résis-
tantes.
L’application
de
ces
mesures
en
1971
s’est
traduite
par
l’interdiction
de
la
tétracycline
en
supplémentation.
En
1972,
un
rapport
similaire
est
publié
aux
Etats-Unis
par
la
Food
and
Drug
Administration
(FDA)
qui
propose,
en
1977,
de
bannir
l’usage
des
pénicillines
et
de
restreindre
celui
des
tétracyclines
comme
promoteurs
de
croissance.
A
ce
jour,
en
raison
de
la
complexité
du
problè-
me,
aucune
décision
n’a
été
prise
(Frap-
paolo
et
Rollins,
1984;
Dupont
et
Steele,
1987).
En
Suède,
une
loi
de
1986
interdit
l’usage
en
tant
qu’additifs
de
tous
les
anti-
biotiques
thérapeutiques.
Dans
le
cadre
de
la
CEE,
depuis
1970
(directive
70-524),
une
réglementation
existe
avec
une
liste
positive
et
des
annexes.

Une
commission
de
l’OMS,
réunie
à
Brême
en
1973,
a
essayé
d’évaluer
les
risques
pour
la
santé
publique
de
l’anti-
biosupplémentation
à
but
zootechnique
et
une
hiérarchie
des
antibiotiques
a
ainsi
été
établie
(Tableau
III)
qui
a
servi
de
base
pour
les
modifications
successives
de
la
directive
70-524.
Dès
1978,
seuls
des
macrolides
et
des
antibiotiques
pré-
sentant
un
risque
plus
faible
restaient
autorisés.
En
France,
où
ces
questions
sont
exa-
minées
par
la
Commission
interministé-
rielle
et
interprofessionnelle
de
l’alimenta-
tion
animale
créée
en
1954,
l’évolution
de
la
législation
a
été
similaire
à
celle
de
la
CEE
et
actuellement
(AM
du
20/11/1987)
restent
seules
autorisées,
parmi
les
anti-
biotiques
à
usage
humain :
la
spiramyci-
ne,
la
virginiamycine
et
la
bacitracine.
L’impact
de
ces
mesures ―
qui
visaient
par
la
réglementation
à
res-
treindre
l’usage
des
antibiotiques
chez
les
animaux
d’élevage
=
sur
la
consomma-
tion
de
ces
molécules
est
difficile
à
appré-
cier.
Braude,
en
1978,
rapporte
que
l’effet
immédiat
du
rapport
Swann
a
été,
en
Grande-Bretagne,
de
réduire
«virtuelle-
ment»
-de
moitié
la
quantité
d’antibio-
tiques
thérapeutiques
utilisés
comme
additifs.
,Cependant
à
partir
de
1974,
leur
usage
s’est
à
nouveau
accru
pour
atteindre,
en
1977,
un
niveau
équivalent
à
celui
atteint
dans
les
années
précédant
le
rapport
Swann.
Apparition
et
principales
étapes
de
l’antibiorésistance
Travaillant
sur
la
pénicilline,
c’est
dès
1940
que
Abraham
et
Chain
observent
que
des
extraits
de
différentes
bactéries
sont
capables
de
détruire
la
molécule.
A
cette
époque
la
pénicilline
n’avait
pas
encore
été
utilisée
en
thérapeutique.
Une
autre
observation
importante
est
faite
par
Mary
Barber
en
1949.
Elle
remarque
que
des
staphylocoques
résis-
tant
à
la
pénicilline
perdent
spontanément
et
à
fréquence
relativement
élevée
l’apti-
tude
à
produire
une
pénicillinase
alors
que
la
réversion
de
ces
souches,
restau-
rant
la
production
de
l’enzyme,
n’a
pas
lieu.
Les
théories
génétiques
de
l’époque,
basées
sur
le
schéma
classique
mutation-
sélection,
n’étaient
pas
satisfaisantes
pour
interpréter
un
tel
phénomène.
Ultérieurement,
l’utilisation
thérapeu-
tique
croissante
d’antibiotiques
apparte-
nant
à
des
familles
de
plus
en
plus
nom-
breuses
conduisit,
en
particulier
chez
les
entérobactéries,
à
l’émergence
de
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
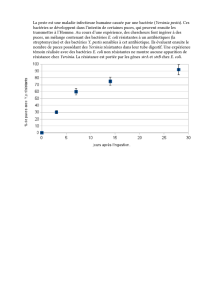
1
/
14
100%