Biologie et potentialités des cellules souches embryonnaires

revue générale
Biologie et potentialités des cellules souches
embryonnaires humaines
Biology and potentialities of human embryonic stem cells
S. Tondeur
1,2,3
S. Assou
1,2,3,5
L. Nadal
1
S. Hamamah
1,2,3,4
J. De Vos
1,2,3
1
CHU Montpellier, Institut de recherche
en biothérapie, Hôpital Saint-Eloi,
Montpellier
2
Inserm U847, Montpellier
3
Université de Montpellier1,
UFR de médecine, Montpellier
4
CHU Montpellier, Unité biologie clinique
d’AMP - DPI, Hôpital Arnaud de
Villeneuve, Montpellier
5
MacoPharma, Tourcoing
Article reçu le 24 novembre 2007,
accepté le 3 mars 2008
Résumé.
Les cellules souches embryonnaires humaines (CSEh) sont des cellu-
les obtenues à partir de la masse cellulaire interne de l’embryon précoce au stade
blastocyste. Dérivées pour la première fois en lignées cellulaires en 1998, elles
peuvent être maintenues en culture, sous forme de colonies indifférenciées, de
manière indéfinie dans certaines conditions. Elles sont caractérisées par deux
propriétés essentielles : pluripotence et auto-renouvellement. Les déterminants
de la pluripotence de ces cellules commencent à être mieux définis, tels que les
facteurs de transcription OCT4 et NANOG, ou les facteurs de croissance FGFb et
IGF2. Les marqueurs caractérisant ces cellules sont des marqueurs de surface
comme SSEA-3, SSEA-4 ou des marqueurs nucléaires, notamment OCT4. Les
CSEh sont capables de se différencier en différents types cellulaires in vitro.
Elles représentent un modèle essentiel et unique dans l’étude du développement
humain précoce et dans le domaine de la médecine régénératrice. Par leur capa-
cité d’auto-renouvellement et leur potentiel de différenciation en de multiples
types cellulaires, les CSEh sont une source illimitée de cellules pour la médecine
régénératrice. Même s’il n’est pas envisageable aujourd’hui pour des raisons
éthiques et scientifiques de les utiliser en clinique, il est primordial d’explorer les
nombreuses potentialités de ces cellules. Ces connaissances pourront immédiate-
ment être appliquées aux modèles de cellules souches adultes et peut-être dans le
futur aboutir à une utilisation en clinique des CSEh.
Mots clés :cellules souches embryonnaires humaines, dérivation, culture,
médecine régénératrice
Abstract.
Human embryonic stem cells (hESC) are obtained from the inner cell
mass from the early embryo at blastocyste stage. Derived in cell lines for the first
time in 1998, they can be maintained in culture in an undifferentiated state
indefinitely under certain conditions. Two essential properties characterize
hESC: pluripotency and self-renewal. Pluripotency is convey by the expression
of specific transcription factors such as OCT4 and NANOG, and is under the
control of growth factors such as IGF2 and FGFb. Markers used to characterize
these cells include surface antigens, notably SSEA-3 and SSEA-4, and nuclear
markers such as OCT4. HESC can differentiate into different cell types in vitro.
They represent a unique and essential model for early human development
research and for regenerative medicine. By their self-renewal capacity and their
potential to differentiate into several cell types, hESC are an unlimited source of
cells enabling to replace or restore lost or damaged cells in numerous diseases.
Even if it is not conceivable today to use them in clinical practice for ethic and
scientific reasons, it seems essential to explore the numerous potentialities of
these cells. This knowledge might be relevant to handle adult stem cells in vitro
and will be mandatory for a therapeutic use of hESC in the future.
Key words
:
human embryonic stem cells, derivation, culture, regenerative medicine
Tirés à part : J. De Vos
abc
Ann Biol Clin 2008 ; 66 (3) : 241-7
doi: 10.1684/abc.2008.0224
Ann Biol Clin, vol. 66, n° 3, mai-juin 2008 241
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Les cellules souches embryonnaires humaines (CSEh)
sont des cellules capables de proliférer in vitro de manière
illimitée dans un état indifférencié et de se différencier en
tous les tissus de l’organisme. Elles sont dites pluripoten-
tes et se distinguent des autres types de cellules souches
humaines : 1) les cellules souches totipotentes, présentes
dans l’embryon de un jour, juste après fécondation,
conduisant au développement d’un être humain ; 2) les
cellules souches multipotentes, déjà engagées vers une
voie de différenciation tissulaire, à l’origine de plusieurs
types de cellules d’un tissu donné, comme les cellules
souches hématopoïétiques médullaires ; 3) les cellules
souches unipotentes, ne donnant naissance qu’à une seule
lignée de cellules comme les cellules souches neurales ou
les kératinocytes. Ces deux derniers types de cellules sont
aussi appelés cellules souches « adultes ».
Origine des CSEh
Les CSEh sont des cellules dérivées in vitro à partir de
l’embryon précoce, le blastocyste, entre le 5
e
et le 6
e
jour
de l’embryogenèse, au stade préimplantatoire c’est-à-dire
avant la nidation dans la paroi utérine. A ce stade le blasto-
cyste a la forme d’une sphère renfermant une cavité liqui-
dienne (blastocèle) et une masse cellulaire interne com-
portant une centaine de cellules (figure 1). In vivo, les
cellules de la masse cellulaire interne vont rapidement
donner naissance aux trois feuillets primordiaux (endo-
derme, mésoderme et ectoderme), puis à tous les tissus
d’un organisme adulte.
Il est possible depuis 1981 chez la souris d’isoler et de
cultiver des cellules souches embryonnaires à partir de la
masse cellulaire interne, tout en maintenant leur capacité
de pluripotence [1]. C’est en 1998 que Thomson et son
équipe isolent pour la première fois des CSEh à partir de
blastocystes humains [2]. Depuis, de nombreuses équipes
ont publié la dérivation de lignées de CSEh. Le taux de
succès de dérivation est très variable et dépend de l’état
(frais ou congelé) des embryons, de leur qualité et de la
technique utilisée. Des embryons à un stade plus précoce
ou plus tardif (au 7
e
-8
e
jour après la fécondation) ont été
également utilisés avec succès. Plusieurs types
d’embryons peuvent être envisagés pour dériver des
CSEh : 1) les embryons congelés obtenus par fécondation
in vitro (FIV), ne faisant plus l’objet d’un projet parental ;
2) les embryons obtenus par FIV mais de qualité insuffi-
sante pour être réimplantés ou congelés ; 3) les embryons
porteurs d’une anomalie génétique recherchée dans le
cadre d’un diagnostic préimplantatoire ; 4) les embryons
créés par « clonage thérapeutique », après transfert d’un
noyau de cellule somatique dans un ovocyte énucléé ;
5) des embryons créés par FIV pour la recherche. La loi
française interdit l’utilisation de ces deux dernières sour-
ces d’embryon.
Pour la dérivation des CSEh, il est recommandé de séparer
la masse cellulaire interne de la couche externe tropho-
blastique. Pour cela, on utilise des techniques d’immuno-
chirurgie (lyse du trophectoderme par réaction
anticorps/complément) ou de séparation mécanique [3].
Une fois isolée, la masse cellulaire interne est mise en
culture dans un milieu contenant du sérum et des facteurs
de croissance, sur une couche de cellules nourricières (fee-
der cells) ou en condition feeder-free, sans cellules de
soutien. Au bout de quelques jours, des colonies de cellu-
les commencent à proliférer autour de la masse cellulaire
interne, générant une lignée de cellules souches embryon-
naires. Une technique permettant la dérivation de CSEh à
partir d’un blastomère unique prélevé sur un embryon de 8
cellules a été décrite récemment [4, 5]. Par cette approche,
on peut théoriquement dériver des CSEh sans destruction
de l’embryon, contournant ainsi en partie les problèmes
éthiques posés par les méthodes dites « destructrices ». Au
1
er
janvier 2006, plus de 400 lignées avaient été dérivées
et publiées (plus ou moins bien caractérisées) dans au
moins 20 pays (essentiellement Etats-Unis, Israël,
Grande-Bretagne, Suède), mais seule une dizaine sont uti-
lisées réellement en recherche. En France, la loi de bioé-
thique du 6 août 2004 a permis aux chercheurs de tra-
vailler sur des lignées de CSEh importées et de débuter les
recherches dans ce domaine dans notre pays. L’autorisa-
tion de dérivation de nouvelles lignées de CSEh a été
accordée à quelques équipes françaises, dont la nôtre,
Figure 1. Embryon humain à J5, obtenu après fécondation in
vitro. On note les cellules pavimenteuses du trophoblaste, et à
l’intérieur de la cavité (le blastocèle), collée aux cellules du
trophoblaste, la masse cellulaire interne qui formera le fœtus. A
ce stade, le blastocyste est entouré par la zone pellucide, une
épaisse membrane translucide constituée de glycoprotéines.
revue générale
Ann Biol Clin, vol. 66, n° 3, mai-juin 2008242
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

depuis l’année dernière. Ces recherches sont très étroite-
ment encadrées par l’Agence de la biomédecine.
Culture et différenciation précoce
des CSEh
Les CSEh sont des cellules de petite taille (2-3 lm) au
rapport nucléo-cytoplasmique élevé, adhérentes, qui for-
ment des colonies plates et compactes en culture
(figure 2). Les techniques de culture de référence sont
similaires à celles utilisées pour les cellules souches
embryonnaires murines (CSEm). Les CSEh ont tout
d’abord été cultivées sur une couche de fibroblastes
embryonnaires murins dont le cycle cellulaire est bloqué
par radiation ou action de composés chimiques (mitomy-
cine C) [2]. Il est également possible de les maintenir et
les amplifier en coculture sur fibroblastes humains prove-
nant par exemple de fragments de peau de prépuce, ou sur
cellules stromales médullaires humaines ce qui permet
d’éviter, dans la perspective d’une éventuelle thérapie cel-
lulaire, la contamination par des éléments pathogènes ou
allergènes transmis par les cellules animales [6]. Le rôle
exact de ces cellules de soutien reste encore mal connu.
Elles fourniraient un signal de communication intercellu-
laire par contact et apporteraient des facteurs solubles
importants pour le maintien de l’état indifférencié des
CSEh, tels que des inhibiteurs de la voie de signalisation
BMP (Bone Morphogenetic Protein). Différents milieux
sont maintenant testés pour une culture sans support cellu-
laire, à base de composants enrichis en matrice extra-
cellulaire tel le Matrigel. Celui-ci est un mélange com-
plexe contenant des molécules extra-cellulaires comme la
laminine, le collagène IV, ainsi que des facteurs de crois-
sance et d’autres substances encore mal définies [7]. A
température ambiante, le Matrigel se solidifie et reconsti-
tue une structure semblable à une membrane basale qui
peut ensuite servir de support à des cellules en culture.
Afin de mieux maîtriser la composition du milieu de cul-
ture, certains utilisent des protéines purifiées comme la
laminine, ou la fibronectine pour recouvrir la surface plas-
tique des plaques de culture. Cette approche est la plus
satisfaisante dans l’objectif d’un milieu de culture parfai-
tement défini et sans protéines animales, mais est très
onéreuse en raison du coût des protéines humaines puri-
fiées. En plus de la matrice qui va servir de support aux
CSEh, la composition du milieu de culture est essentielle.
Le milieu de référence initialement décrit contenait du
sérum de veau fœtal (SVF). En raison de la grande varia-
bilité des lots de SVF, dont beaucoup étaient inadaptés à la
culture des CSEh, la plupart des équipes utilisent doréna-
vant le KnockOut Serum Replacement (KO-SR). Les fac-
teurs de croissance et cytokines nécessaires aux CSEh
diffèrent de ceux indispensables au maintien en culture
des cellules souches embryonnaires murines. En effet,
chez l’homme, au contraire du modèle murin, le LIF n’est
pas requis pour la culture des CSEh [2]. En revanche, le
basic Fibroblast Growth Factor (bFGF ou FGF-2) est
nécessaire à la culture des CSEh in vitro et est le facteur
critique de la plupart des protocoles de culture de ces
cellules. Récemment Bendall et al. ont montré le rôle
important joué par un autre facteur de croissance, l’Insulin
Growth Factor 2 (IGF2) dans la prolifération et la survie
des CSEh [8]. Certains protocoles utilisent du milieu
« conditionné », surnageant de culture de fibroblastes [9].
Tous ces milieux et ces techniques de culture restent
encore très hétérogènes et sont difficiles à standardiser. De
nombreux travaux sont publiés pour déterminer le milieu
de culture optimal et le mieux caractérisé possible (com-
position en protéines connue) pour une culture à long
terme [10-12]. L’obtention de conditions de culture stan-
dardisées est indispensable pour l’utilisation de ces cellu-
les en recherche. Pour pouvoir utiliser les CSEh ou leurs
dérivés en thérapie cellulaire, il sera en effet nécessaire
AB
Figure 2. Colonie de cellules souches embryonnaires humaines indifférenciées (flèche pleine) sur fibroblastes de soutien (flèche en
pointillé) (microscope optique x 20 (A) et x 50 (B)). Les cellules souches embryonnaires humaines sont des cellules de petite taille
formant des colonies plates et très compactes en culture.
Cellules souches embryonnaires humaines
Ann Biol Clin, vol. 66, n° 3, mai-juin 2008 243
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

d’éliminer de nos méthodes de culture les contaminations
potentielles par des pathogènes ou par des protéines ani-
males.
Afin d’éviter la différenciation spontanée des cellules sou-
ches, il est important de renouveler quotidiennement le
milieu de culture et de diviser les colonies avant qu’elles
n’arrivent à confluence. Différentes techniques de repi-
quage existent : séparation enzymatique (dissociation par
la trypsine, la collagénase, la dispase...) ou séparation
mécanique. Il est possible d’effectuer ainsi jusqu’à 150
passages soit, à raison de4à8passages par mois, de les
maintenir en culture pendant 1 à 3 ans. Au cours du temps
une adaptation des cellules peut survenir avec des change-
ments phénotypiques et génotypiques [13]. Il est donc
nécessaire de s’assurer que les CSEh garderont sur le long
terme leur phénotype immature, un caryotype normal
et leur capacité à se différencier. Les remaniements
chromosomiques pouvant survenir après culture au long
cours sont le plus fréquemment une trisomie 12 ou une
trisomie 17. Ces anomalies surviennent notamment après
action de la trypsine qui dissocie les colonies, les cellules
se retrouvant alors sous forme isolée, avec possible pres-
sion de sélection positive pour les cellules ayant un avan-
tage de survie et/ou de prolifération du fait de l’anomalie
génétique.
Les CSEh sont caractérisées in vitro et in vivo par leur
potentiel à donner naissance aux 3 feuillets primordiaux
embryonnaires : endoderme, ectoderme, mésoderme puis
tous les tissus adultes. Cultivées en l’absence des facteurs
nécessaires au maintien de leur état indifférencié, en
conditions non adhérentes, les cellules vont former des
agrégats qui se différencient spontanément de manière
non organisée en structures vésiculaires appelées « corps
embryoïdes ». Au sein de ces corps embryoïdes sont
détectés des marqueurs de l’ectoderme (tissu neurologi-
que, par exemple bIII tubuline, NCAM1), du mésoderme
(tissu cardiaque, par exemple actine du muscle lisse,
VEGFR2), et de l’endoderme (tissu digestif, respiratoire,
par exemple alpha-fœtoprotéine) [14, 15]. Le caractère
prolifératif et la propriété de pluripotence sont démontrés
également in vivo après injection des CSEh chez la souris
immunodéficiente (en sous-cutané, intra-rénal ou intra-
testiculaire). Les CSEh injectées vont générer des térato-
mes solides comportant des cellules des 3 couches primai-
res et des cellules indifférenciées [2].
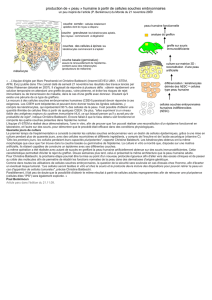
Déterminants de la pluripotence
des CSEh
Pluripotence et auto-renouvellement sont les caractéristi-
ques majeures des CSEh. La pluripotence des CSEh est
sous le contrôle de réseaux de facteurs de transcription et
de modifications épigénétiques (facteurs intrinsèques), et
sous le contrôle de l’environnement cellulaire (facteurs
extrinsèques) (figure 3).
Déterminants intrinsèques
A ce jour, seul un nombre limité de gènes participant à la
propriété de pluripotence des CSEh a été identifié, parmi
lesquels figurent Oct4, Fgf4, Foxd3, LeftyA, Nanog et
Sox2. Oct4 (Oct3/4, POU5F1) est le marqueur le plus
utilisé. Ce facteur de transcription de la famille POU
(PICT/OCT/UNC) joue un rôle central dans le développe-
ment des CSEh et le maintien de la pluripotence [16]. Il
agit en collaboration avec Sox2, de la famille Sox et avec
Nanog, un autre facteur de transcription à homéodomaine.
Ces 3 facteurs se fixent sur les promoteurs d’un groupe de
gènes exprimés dans les CSEh, codant pour des molécules
impliquées dans le maintien de la pluripotence (incluant
NANOG et Oct4 eux-mêmes) [17]. Ils sont indispensables
à l’auto-renouvellement des CSEh et sont retrouvés forte-
ment exprimés dans ces cellules, leur taux d’expression
diminuant au cours de la différenciation. La disparition de
Oct4, mise en évidence par RT-PCR ou par immunofluo-
rescence, signe la perte de pluripotence des CSEh et donc
leur différenciation.
La structure chromatinienne des CSEh semble jouer un
rôle particulièrement important dans la pluripotence des
CSEh. Les caractéristiques épigénétiques, comme la
méthylation de l’ADN ou l’acétylation et la méthylation
des histones influencent la capacité de pluripotence des
cellules. L’ADN de la région promotrice du gène Oct4 est
déméthylé, donc sous forme active dans les CSEh. La
méthylation de cette région empêche l’expression d’Oct4
et induit la différenciation des cellules. De manière réver-
sible, la déméthylation après exposition à un agent démé-
thylant, la 5-aza-2-deoxycytidine, ou la déacétylation des
histones par la trichostatine A de corps embryoïdes murins
induit une dédifférenciation vers des cellules de phéno-
type immature [18]. Plusieurs études récentes montrent
TGFb
ACTIVIN
NODAL
bFGF
NOGGIN
BMP
OCT4
NANOG
SOX2
ßCATENIN
PLURIPOTENCE
WNT
Figure 3. Schéma des différents facteurs impliqués dans la pluri-
potence des cellules souches embryonnaires humaines.
revue générale
Ann Biol Clin, vol. 66, n° 3, mai-juin 2008244
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

que l’épigénome des CSEh est particulièrement original,
en accord avec la notion plus ancienne d’une mise à plat
complète des modifications de la chromatine au cours du
développement embryonnaire précoce [19].
Les déterminants intrinsèques de la pluripotence sont
associés à des modifications épigénétiques majeures et
sont aussi importants pour activer un programme
d’expression génique spécifique des CSEh que pour répri-
mer des programmes de différenciation.
Déterminants extrinsèques
La culture à long terme des CSEh sous forme indifféren-
ciée et pluripotente nécessite certains facteurs de crois-
sance ou cytokines exogènes, notamment le bFGF.
D’autres facteurs indispensables sont apportés par les cel-
lules fibroblastiques de soutien. Des modifications mineu-
res des conditions de culture peuvent induire rapidement
la différenciation des CSEh. Ces facteurs extrinsèques
comprennent les interactions intercellulaires, des facteurs
diffusibles dans le milieu de culture et la matrice extra-
cellulaire.
Les CSEh poussent en colonies compactes et sont diffici-
les à cloner. Les contacts intercellulaires entre CSEh font
intervenir des gap junctions. Les CSEh expriment 18 des
20 connexines qui constituent les gap junctions, en parti-
culier les connexines 43 et 45. Cette forte expression se
traduit par une intense interconnexion entre les cellules
d’une colonie avec diffusion rapide de marqueurs intra-
cellulaires d’une cellule à une autre au sein de la colonie
[20]. De nombreuses voies de signalisation intracellulaires
sont impliquées dans le maintien de l’état indifférencié
des CSEh, notamment la voie TGFb/activin/nodal, per-
mettant une forte expression de SMAD2/3 [21]. Au
contraire, l’activation de la voie des BMP entraîne la dis-
parition d’Oct4 et induit la différenciation des CSEh. La
suppression des BMP par des antagonistes comme noggin
ou gremlin sécrétés par les fibroblastes de soutien est
nécessaire à la pluripotence [22]. Les membres de la
famille WNT semblent également importants, via la
dégradation de la bêta-catenin par la glycogène synthase
kinase-3. Bien que le principe de l’activation de la voie
WNT soit repris dans certains protocoles de culture des
CSEh, l’activation de cette voie de signalisation n’est pas
suffisante. Le rôle exact de la voie WNT reste discuté [23].
L’auto-renouvellement et la pluripotence des CSEh
dépendent d’une balance finement régulée entre des molé-
cules de signalisation nombreuses et variées, le rôle précis
de chacune restant à déterminer.
L’évaluation du caractère indifférencié et pluripotent des
CSEh peut se faire par la mise en évidence en immuno-
fluorescence d’un panel de marqueurs membranaires
comme les antigènes Stage-Specific antigens SSEA-3,
SSEA-4 (absents chez la souris chez laquelle on trouve
SSEA-1), les glycoprotéines tumor-recognition antigen
TRA-1-60 et TRA-1-81 (Ac monoclonaux dirigés contre
des protéoglycanes kératanes sulfates), Thy1, et des phos-
phatases alcalines ou ALPL (alkaline phosphatase
liver/bone/kidney). La présence du marqueur Oct4 est
mise en évidence par immunofluorescence ou par techni-
ques de biologie moléculaire. L’étude du caryotype des
CSEh est indispensable pour s’assurer du caractère non
transformé des cellules tout au long de la culture.
Potentialités et intérêt des CSEh
Les cellules souches embryonnaires représentent une
source abondante et renouvelable à l’infini de cellules
fonctionnelles pour de nombreuses applications en recher-
che fondamentale et en thérapeutique.
Les lignées de CSEh fournissent de nouveaux outils pour
l’étude des mécanismes moléculaires mis en jeu au cours
du développement humain normal, qui ne peuvent, pour
des raisons éthiques évidentes, être étudiés in utero. Elles
permettront de mieux appréhender la formation normale
des tissus à partir du stade embryonnaire, de comprendre
le développement des organes et également d’explorer
l’oncogenèse et le développement des tumeurs d’origine
embryonnaire.
En théorie, de par leur capacité d’auto-renouvellement, les
CSEh pourraient générer un nombre illimité de cellules
différenciées spécialisées, fournissant une source de cellu-
les très intéressante pour la thérapie cellulaire. Pluripo-
tence et prolifération sont deux propriétés précieuses pour
la production de cellules pour une médecine régénératrice.
A plus ou moins long terme, et sous réserve d’une modifi-
cation de la loi, les CSEh pourraient être utilisées en théra-
peutique pour traiter des pathologies aujourd’hui incura-
bles. En 2004 il était estimé qu’une application de thérapie
cellulaire à partir de CSEh mettrait cinq ans pour passer
du laboratoire à une utilisation clinique validée par des
essais cliniques [24]. La mise au point de conditions de
dérivation compatibles avec les bonnes pratiques de la
thérapie cellulaire est un préalable indispensable à cette
perspective. Les CSEh et leurs dérivés semblent avoir des
propriétés immunogéniques faibles in vivo [25]. Ce privi-
lège immun permet d’envisager les CSEh comme un
espoir pour les patients qui n’auraient pas de donneur
HLA compatible.
In vitro il a été montré que des CSEh pouvaient se diffé-
rencier en cellules fonctionnelles de nombreux tissus : cel-
lules hépatiques, pancréatiques, cardiaques, neuronales,
hématopoïétiques... Le diabète de type I représente une
des pathologies qui pourrait fortement bénéficier de la
thérapie de remplacement cellulaire. Chez le patient dia-
bétique, les îlots de Langerhans du pancréas sont détruits
Cellules souches embryonnaires humaines
Ann Biol Clin, vol. 66, n° 3, mai-juin 2008 245
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.
 6
7
6
7
1
/
7
100%