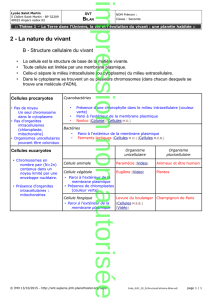
Transport du réseau trans-Golgien à l`extérieur de la cellule

TRAFIC DES VÉSICULES
DANS LA CELLULE

2
TRAFIC VÉSICULAIRE INTRA-CELLULAIRE
I. Les mécanismes moléculaires du transport par
membrane et le maintien de la diversité des
compartiments
II. Transport à partir du reticulum endoplasmique à
travers le Golgi
III. Transport du TGN vers les lysosomes
IV. Transport depuis la membrane plasmique vers
l'intérieur de la cellule : endocytose
V. Transport du TGN vers l'extérieur de la cellule :
exocytose

3
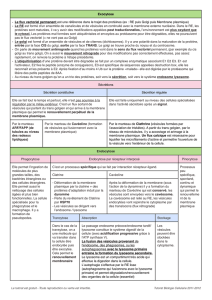
V - EXOCYTOSE

4
Schéma du routage

5
Introduction
•On a vu le système digestif interne de la
cellule qui se termine dans le lysosome
•On revient au Golgi dont les voies se
terminent à l'extérieur de la cellule
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
1
/
52
100%