L`évolution du squelette

Evolution du squelette
Dès l'apparition des animaux pluricellulaires, le squelette fut nécessaire. Par son rôle de
soutien, il permet de lutter efficacement contre la gravité et de maintenir la cohérence de
l'organisme. Ce squelette est généralement constitué de parties dures mais peut aussi bien être
de composition liquide (qui, rappelons-le sont incompressibles). Il va prendre de l'importance
dans la conquête du milieu aérien par les animaux.
Chez les Unicellulaires
Nous ne pouvons pas parler de squelette sans citer le cytosquelette des cellules. Toute cellule
Eucaryote, animale ou végétale, possède des molécules dont le rôle est de contrôler la
mobilité des organites et de la cellule elle-même, ainsi que de donner une forme spécifique à
celle-ci pouvant améliorer sa fonction.
Ainsi les Protozoaires, Flagellés, Ciliés, Acanthaires, Rhizopodes peuvent se déplacer grâce à
l'action de ce cytosquelette. Certains, comme les Foraminifères, les Radiolaires, possèdent un
test. C'est un exosquelette calcaire ou siliceux produit par la cellule. De nombreuses roches
sont constituées par l'accumulation de ces coquilles sur les fonds marins. Ainsi la craie est
formée par le dépots des concrétions calcaires protégeant une algue, la coccolithophoride. Le
fond des océans n'est recouvert que par les tests de Radiolaires car leur composition siliceuse
les protégent de la dissolution à ces profondeurs.
Chez les Invertébrés
Un premier squelette
Dès les Spongiaires on trouve un squelette. Certes, celui-ci est diffus; ce sont des spicules
disséminés dans les tissus mais sa présence est nécessaire à la bonne tenue de l'animal. Ces
spicules, présents dans la mésoglée, sont secrétés par des cellules ectodermiques, les
scléroblastes. Ils sont d'origine minérale (calcaire ou siliceuse). Ils peuvent être secondés par
la spongine, une protéine fibreuse, sorte de collagène, qui fournit à l'éponge des propriétés
d'élasticité et de souplesse que l'Homme a su exploiter. La mésoglée sert de support aux
feuillets cellulaires et intervient donc dans une moindre proportion comme tissu de soutien.
Certains formes fixées de Cnidaires, les polypes, possèdent une gaine externe d'origine
ectodermique, le périderme, qui joue le rôle de squelette externe. Mais c'est au niveau des
Octocoralliaires ou Coraux que l'on peut observer un squelette plus abouti. Ce sont des formes
coloniales enchassées dans une mésoglée commune. Chaque polype produit des spicules, par
des cellules ectodermiques, qui vont fusionner et former un véritable squelette souvent
recherché par l'Homme pour ses formes et ses couleurs.

La forte proportion du corps en eau de ces organismes, notamment les formes libres ou
méduses, compense l'absence d'un tissu de soutien.
Le squelette qui n'en est pas un
Avec les Plathelminthes et l'apparition concomitante du mésoderme, le squelette "en dur"
n'est plus vraiment nécessaire. C'est le parenchyme, d'origine mésodermique, qui va
remplacer avec beaucoup plus d'efficacité le rôle de la mésoglée. Le tissu s'insinue entre les
différents organes et jouer le rôle de squelette hydrostatique.
Un tégument fin recouvre les formes de Plathelminthes parasites (Trématodes et Cestodes). Il
a pour rôle principal de protéger l'animal contre l'acidité du tube digestif dans lequel se trouve
le parasite. Il complète le rôle du parenchyme en augmentant la résistance aux pressions
externes.
La formation d'une cavité générale chez les Némathelminthes a considérablement développé
l'importance du squelette pour maintenir l'intégrité de l'organisme. C'est le liquide remplissant
cette cavité, le pseudocoelome, (reste du blastocoele embryonnaire) qui joue le rôle de
squelette hydrostatique. Un tégument épais mais élastique permet aux fluides internes
d'exercer la pression nécessaire à leur rôle de soutien.
Cette disposition est améliorée chez les Annelides avec la formation du coelome, une cavité
délimité par un tissu mésodermique. Le fait que ce coelome soit limité, et qu'il y ait une
segmentation en plusieurs sacs coelomique augmente considérablement l'efficacité dans son
rôle de soutien. Le tégument est d'ailleurs bien plus fin que chez les Némathelminthes
(excepté pour les tubicoles). De plus le coelome intervient dans beaucoup d'autres fonctions
de l'organisme en permettant un meilleur développement des organes, leur disposition
métamérique, la formation de nouvelles structures (vaisseaux sanguins). C'est pourquoi ce
dispositif anatomique a été conservé tout au long de l'évolution.
L'exosquelette
C'est le tégument qui va toutefois prendre de l'importance chez les Invertébrés supérieurs. Son
rôle prédominant commence à apparaître avec les Mollusques. Ces animaux secrètent en effet
une coquille au niveau de leur manteau. Bien que ne recouvrant pas complètement le corps
cette coquille leur sert de protection. Cet exosquelette est d'origine ectodermique et formé de
calcaire. Le manteau produit la nacre (chez les huîtres une impureté à ce niveau provoque la
formation d'une perle) tandis qu'une zone de la cavité palléale (le bourrelet palléal) permet
l'augmentation en taille. Cette coquille étant facilement fossilisable est un critère essentiel de
détermination des mollusques. Sa présence dans les roches permet de les dater rapidement.
Les Mollusques terrestres et pélagiques (nageurs) montrent un allégement important de ce
squelette. Ce n'est que chez les Céphalopodes que l'on observe une régression importante de
la coquille. Celle-ci devient interne et s'allége considérablement (os de seiches, plumes de
calmar).
On peut également citer le pseudocrâne des Céphalopodes qui protégent leur cerveau.
Tous ces organismes sont encore majoritairement aquatiques. De toutes façons ils dépendent
de la présence d'eau ou du moins d'une forte humidité. Avec les Arthropodes cette
dépendance n'a plus lieu. C'est grâce à leur tégument que ces animaux ont pu s'affranchir des

deux contraintes les plus importantes du milieu aérien : la gravité d'une part et la
déshydratation de l'autre. En milieu aquatique le corps est soutenu par l'eau, en milieu aérien
ce n'est pas le cas. La cuticule des Arthropodes épousant parfaitement le corps est légère (il
n'y a quasiment pas de formations minérales, sauf pour les crustacés qui sont aquatiques) et
imperméable tout en étant souple et résistante ! Ce tégument est pluristratifié, c'est à dire
composé de plusieurs couches, toutes produites par l'épiderme. Au niveau des articulations et
entre les segments la cuticule présente des repli sous-cutané qui augmentent la souplesse du
tégument en autorisant la variation du volume du corps. La présence d'un tel exosquelette
nécessite des mues pour permettre la croissance de l'animal. A chaque mue une grande partie
de la cuticule est dégradée et recyclée par l'individu ce qui représente une réelle économie par
rapport aux autres dispositifs. Quand la nouvelle cuticule est formée l'étape du tannage est
nécessaire pour le durcissement de la cuticule.
Chez les espèces aériennes la cuticule recouvre les trachées jusqu'au plus profond de
l'organisme.
Le pagure ou Bernard l'hermitte, crustacé à corps mou, vit dans une coquille de Gastéropode
abandonné pour compenser le manque de carapace.
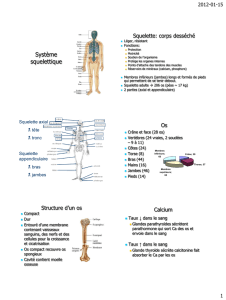
Chez les Vertébrés
Le squelette des Vertébrés est interne. Il est composé de plusieurs pièces solides articulées
entre elles. Mais il ne faut pas oublier que les phanères (productions cutanées de l'épiderme,
comme les poils) ont pu former un véritable exosquelette. Ainsi la tortue est pourvue d'une
carapace formée par, d'une part , les écailles épidermiques (propres aux reptiles), d'autre part,
des os dermiques juxtaposés qui sont en relation avec le squelette interne. On retrouve cette
disposition anatomique chez certains Dinosaures, les Ankylosaures (Crétacé), et des
Mammifères actuels, les Tatous.
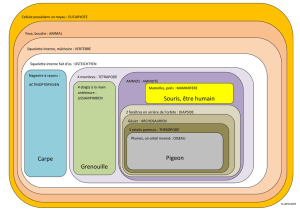
D'un point de vue anatomique on peut facilement distinguer trois régions dans le squelette
interne :
- le squelette céphalique, ou crâne,
- le squelette axial, qui correspond la plupart du temps à la colonne vertébrale,
- le squelette appendiculaire, qui compose les membres et est associé au squelette axial
par l'intermédiaire du squelette zonal (les ceintures).
Le squelette céphalique
Le crâne est constitué par deux régions distinctes d'origines différentes. Le Neurocrâne qui
protége le cerveau et le splanchnocrâne qui constitue le squelette viscéral et formera la
machoire.
Le Neurocrâne
Chez les Agnathes ce n'est qu'une plaque cartilagineuse repliée sur les cotés et possédant deux
capsules otiques dans la partie postérieure. Cette disposition se complique chez les Sélaciens
où l'on peut observer un paléocrâne (proche de celui des Agnathes) et l'apparition d'un
néocrâne. Le paléocrâne englobe partiellement les capsules olfactives et otiques (les capsules
optiques restent indépendantes). Le néocrâne correspond à la fusion de territoires vertébraux
pour compléter et agrandir le crâne dans sa partie postérieure.

Le crâne s'ossifie et il se forme des fosses temporales permettant de l'alléger. En fonction du
nombre et de la disposition de ces fosses on distingue :
- les Anapsides : ils n'ont pas de fosses, ce sont les Amphibiens et les Chéloniens.
- les Euryapsides : Ils possèdent une fosse temporale dans la partie postérieure haute du crâne.
- les Diapsides : Ils possèdent deux fosses temporales (Crocodiliens) qui peuvent fusionner
avec les orbites (Oiseaux).
- les Synapsides : Ils possèdent une fosse temporale dans la partie postérieure basse du crâne
(Reptiles mammaliens, Mammifères).
Le neurocrâne, dès les Amphibiens, migre peu à peu vers l'arrière ce qui permet au museau de
s'individualiser. La capsule otique se rapproche de l'articulation de la machoire (ce qui
permettra d'une part l'intégration de certains os dans l'oreille et d'autre part l'agrandissement
du crâne latéralement).
Le Splanchnocrâne
C'est lui qui soutient les fentes branchiales du pharynx. Il va rapidement évoluer pour former
la machoire. Les fentes branchiales (en position intermétamériques) sont soutenues par la
présence d'arcs situés de part et d'autres du tube digestif qui se rejoignent au milieu du ventre.
Chaque arc provient de l'évolution d'un segment mésodermique de l'embryon. Ils sont
généralement au nombre de 7 paires, un par fente.
- L'arc le plus antérieur permet de soutenir les parois de la bouche. Il est constitué de deux
cartilages. A partir des Gnathostomes, un forme la base du crâne (cartilage palato-pterygo-
carré), l'autre forme la machoire inférieure (cartilage de Meckel). C'est l'arc Mandibulaire. Il
limite la première fente branchiale, l'évent. Il faut toutefois signaler l'existence d'un arc
prémandibulaire qui donne les trabécules, os incorporés au neurocrâne et permettant son
agrandissement vers l'avant.
- Le deuxième arc intervient dans la suspension de cette machoire inférieure au crâne. Seule
sa partie dorsale joue un rôle, c'est le hyomandibulaire. La partie ventrale, l'hyoïde, reste libre.
C'est l'arc Hyoïde. Il limite la seconde fente branchiale.
- Viennent ensuite des arcs plus classique qui limitent les branchies. On en compte 5.
Au cours de l'évolution les deux premiers arcs vont se transformer progressivement en
mâchoire. Les arcs branchiaux quant à eux régressent chez les Tétrapodes. Leurs résidus
participent au squelette de la langue des Reptiles et Oiseaux et aux cartilages du pharynx chez
les mammifères. L'absence de branchies permet toutefois l'apparition du cou, donc d'une

meilleure mobilité de la tête et d'une meilleure appréciation de l'environnement extérieur.
Ces arcs ne restent cartilagineux que chez les Chondrichtyens, ils deviennent osseux avec les
Ostéichtyens et sont complété par des os de membranes.
Le palato-pterygo-carré va donner trois os indépendant : le palatin, le pteroïde et le carré. Ils
se soudent au neurocrâne chez les Poissons pour former la cavité buccale (palais). Le cartilage
de Meckel s'ossifie dans sa partie antérieure et donne l'articulaire. Le reste est remplacé par
des os de membranes qui forment la mâchoire inférieure (l'os terminant la mâchoire est appelé
le dentaire). Le mode d'articulation est alors de trois types en fonction de l'évolution :
- La suspension Amphistylique, chez les Sélaciens primitifs. Le palatin et le carré forment
deux points de soudures (ou processus) de la machoire supérieure avec le neurocrâne.
l'hyomandibulaire est lié au carré ou libre.
- La suspension hyostylique chez les Poissons en général (excepté les holocéphales). dans ce
cas seule la partie antérieure de la machoire supérieure (palatin) est fortement soudée au
neurocrâne. C'est l'hyomandibulaire qui retient le carré.
- La suspension autostylique chez les Tétrapodes, dans ce cas une expansion du ptéroïde
renforce la soudure de la machoire au crâne. L'articulation se fait aussi principalement avec le
carré, l'hyomandibulaire régresse et forme la columelle dans la capsule otique.
La première fente branchiale, l'évent, n'apparait que chez les Sélaciens et les Chondrostéens,
de plus chez les Ostéichtyens un opercule osseux, fixé sur l'hyoïde, recouvre les arcs
branchiaux. Jusqu'aux Reptiles l'articulation de la machoire se fait entre le carré et
l'articulaire. L'hyomandibulaire lui forme une petite baguette qui entre dans le système auditif
des Amphibiens, il transmet les vibrations extérieures au crâne. Puis il forme un sorte
d'anneau léger, l'étrier, qui s'incorpore dans l'oreille des Reptiles. La réduction du museau
chez les mammifères leur permet d'avancer l'articulation de la machoire. Ainsi le carré et
l'articulaire, libérés de la machoire règressent et forment respectivement le marteau et
l'enclume de l'oreille moyenne. La machoire inférieure n'est plus composée que d'un os, le
dentaire, articulé avec le squamosal un os descendu du neurocrâne. C'est l'articulation
mammalienne. L'articulation reptilienne se retrouve donc entre marteau et enclume, dans
l'oreille !
A partir des Reptiles (crocodiliens), le palais est doublé par un palais secondaire qui permet
l'indépendance entre la bouche et la cavité nasale. Il est désormais possible de manger
(mastiquer) et respirer en même temps !
Le squelette axial
Le premier squelette axial est la chorde. Présente en totalité chez les Agnathes, elle régresse
au fur et à mesure de l'évolution, jusqu'à disparaître (elle reste visible chez les embryons). Elle
est remplacé par un squelette axial secondaire composé de vertèbres.
Chez les Poissons on peut déjà observer une certaine régression. Ainsi la chorde des Sélaciens
présentent un pincement au niveau de chaque vertèbre. Au niveau des Amphibiens elle est soit
restreinte à être en position intervertébrale soit présente au centre de la vertèbre. Chez les
jeunes Mammifères elle compose les disques intervertébraux mais disparaît chez l'adulte.
Les vertèbres sont composées de plusieurs pièces qui s'appliquent contre la chorde :
- deux pièces dorsales forment un arc en délimitant le canal rachidien qui contient la moelle
épinière. C'est l'arc neural.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%