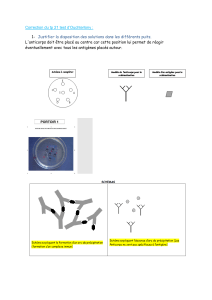
Immunologie La boite à outils des immunologistess





 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%