Implication des phagocytes mononucléés dans les pathologies

UNIVERSITE PARIS VI – PIERRE ET MARIE CURIE
ECOLE DOCTORALE DE PHYSIOLOGIE ET PHYSIOPATHOLOGIE
THESE DE DOCTORAT
Spécialité de Physiologie et Physiopathologie
Présentée par John PIRAULT
Implication des phagocytes mononucléés dans les
pathologies immuno-inflammatoires chroniques :
exploration d’un modèle murin d’athérosclérose
et le xanthogranulome chez l’homme.
Thèse dirigée par Philippe GIRAL et Philippe LESNIK
Soutenue le 02 Février 2012
Jury :
Dr Giuseppina CALIGIURI
rapporteur
Dr Giulia CHINETTI
rapporteur
Dr Carole ELBIM
examinateur
Dr Muriel LAFFARGUE
examinateur
Dr Pierre AUCOUTURIER
président
Dr Philippe GIRAL
directeur
Dr Philippe LESNIK
directeur
tel-00833457, version 1 - 12 Jun 2013

2
Remerciements
Je tiens tout d’abord à remercier Giuseppina Caligiuri, Giulia Chinetti, Muriel Laffargue,
Carole Elbim et Pierre Aucouturier qui représentent l’ensemble des membres du jury qui a
accepté de réviser ce travail de thèse.
Mes remerciements s’adressent aussi à l’ensemble de l’unité INSERM UMR_S 939.
A John Chapman pour m’avoir accueilli en octobre 2008 dans l’unité qu’il dirigeait.
A Philippe Giral pour avoir accepté d’être mon directeur de thèse, pour m’avoir apporté ses
connaissances cliniques et sa vision de la collaboration entre cliniciens et chercheurs.
A Philippe Lesnik, co directeur de ma thèse, qui m’a suivi quotidiennement au cours de ce
parcours doctoral. Merci pour l’ensemble de ton soutient dans l’orientation et le choix des
projets qui ont fait partie de cette thèse. Merci pour l’apport de tes connaissances scientifiques
pratiques et administratives qui ont bordé ces années. Merci à l’ensemble des acteurs qui ont
participé à ces travaux parmi lesquels Laura Bouchareychas, Wilfried Le Goff, Maryse
Guerin, Flora Saint Charles, Raphaël Szalat.
Enfin, merci à ma famille, mes parents Brigitte et Alain et mon frère Joffrey pour leur soutien
quotidien dans l’établissement de ma vie personnelle et professionnelle.
tel-00833457, version 1 - 12 Jun 2013

3
Contenu
Remerciements...........................................................................................................................2
Contenu ......................................................................................................................................3
Abbréviations .............................................................................................................................4
Avant-propos..............................................................................................................................6
I. L’athérosclérose. ................................................................................................................8
A. Généralités......................................................................................................................8
1. Classification et complications...................................................................................8
2. Facteurs de risques. ..................................................................................................11
B. Aspects cellulaires et moléculaires de l’athérosclérose. ..............................................11
1. A l’origine la dysfonction endothéliale....................................................................11
2. L’impact des dyslipidémies : rôle pro-athérogène des LDL et de
l’hypercholestérolémie.....................................................................................................12
3. Inflammation et apoptose.........................................................................................14
II. Le système immunitaire : acteur pro et anti-athérogène. .................................................26
A. L’immunité adaptative, lignée lymphoïde. ..................................................................27
1. Les lymphocytes associés à l’augmentation de l’athérosclérose..............................29
2. Les lymphocytes associés à la protection de l’athérosclérose..................................36
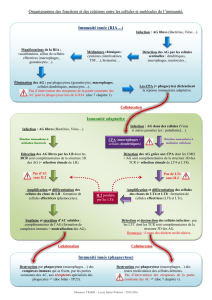
B. L’immunité innée, lignée myéloïde. ............................................................................44
1. Les monocytes : précurseurs myéloïdes et senseurs circulants................................45
2. Les cellules dendritiques, interface entre l’immunité innée et adaptative. ..............48
3. Les macrophages, phagocytes majeurs impliqués dans l’athérosclérose.................54
III. Objectifs .......................................................................................................................63
IV. Exposé des travaux de thèse.........................................................................................66
1. Article 1: Increasing survival of macrophages exerts protective functions on
progression of established atherosclerotic lesions, through decreasing plasma cholesterol
levels, and dampening inflammation. ..............................................................................66
2. Article 2: Impaired macrophage lipid homeostasis and imbalance in dendritic cell
subsets are key features in normolipemic xanthoma associated with monoclonal
immunoglobulin. ..............................................................................................................93
Discussion générale................................................................................................................126
1. Discussion sur la survie des macrophages dans l’athérosclérose murine. .............126
2. Discussion sur l’étude d’une pathologie humaine : le xanthogranulome...............137
Références..............................................................................................................................145
Annexes..................................................................................................................................161
tel-00833457, version 1 - 12 Jun 2013

4
Abbréviations
ABCA1/G1 ABC transporter A1/G1
AIM apoptosis inhibitor of macrophage
Apoe apolipoprotein E
BCR B cell receptor
BDCA-2 blood dendritic cell antigen 2
CCR2 chemokine (CC motif) receptor 2
CD cellule dendritique
CX3CR1 chemokine (C-X3-C motif) receptor 1
DAMP damage associated molecular pattern
DD death domain
DISC death inducing signalling complex
DT diphtheria toxin
FasL Fas ligand
GM-CSF granulocyte macrophage colony stimulating factor
HDL high density lipoprotein
Hsp heat shock protein
ICAM1 intercellular cell adhesion molecule 1
IFN interferon
IgG/M… immunoglobulin G/M…
IL-6/10… interleukine 6/10…
LDL low density lipoprotein
LDLox LDL oxydée
tel-00833457, version 1 - 12 Jun 2013

5
LXR liver X receptor
M-CSF macrophage colony stimulating factor
MDA-LDL malondialdehyde-LDL
MerTK Mer tyrosine kinase
mPDCA-1 mouse plasmacytoid dendritic cell antigen 1
NFκB nuclear factor kappa b
NLR NOD-like receptor
PAMP pathogen associated molecular pattern
PC phosphorylcholine
pDC plasmacytoid dendritic cell
PI3K phosphoinositide 3 kinase
PPAR peroxisome proliferator activated receptor
PRR pattern recognition receptor
PTX pentoxyfilin
RORγt retinoid related orphan receptor γ t
SCID severe combined immunodeficiency
SOCS suppressor of cytokine signalling
STAT signal transducer and activator of transcription
TCR T cell receptor
TGFβ transforming growth factor
TLR Toll-like receptor
TNF tumor necrosis factor
VCAM1 vascular cell adhesion molecule 1
tel-00833457, version 1 - 12 Jun 2013
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
1
/
224
100%








![Poster CIMNA journée CHOISIR [PPT - 8 Mo ]](http://s1.studylibfr.com/store/data/003496163_1-211ccc570e9e2c72f5d6b6c5d46b9530-300x300.png)