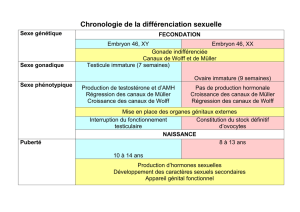
Le canal de Wolff induit la formation de rostium

/. Embryol. exp. Morph. Vol. 25, 1, pp. 115-129, 1971
Printed in
Great
Britain
Le canal de Wolff induit la
formation de rostium mullerien: demonstration
experimentale chez Tembryon de Poulet
Par ELIANE DIDIER1
Laboratoire de Biologic animate, Universite de Clermont-Ferrand
(Directeur: Professeur Lutz)
RESUME
L'analyse experimentale du role du canal de Wolff dans la formation de l'ostium du canal
de Muller a ete entreprise par blocage du canal de Wolff
en
avant et a differents niveaux de la
zone presomptive ostiale.
Si le blocage du canal de Wolff a lieu entre les somites 13 a 19, l'ostium se forme toujours
a cote du secteur du canal de Wolff edifie.
Si le blocage a lieu entre les somites 9 a 13, l'agenesie ostiale et mullerienne est absolue.
On en conclut que la zone presomptive de l'ostium mullerien est situee entre les somites
.13 a 19 chez l'embryon de Poulet de 1 a 1,5 jour, et que la differentiation ostiale necessite
la presence du canal de Wolff au contact de ce vaste territoire presomptif.
INTRODUCTION
Alors que les premieres etapes de la formation du canal de Miiller de
l'embryon de Poulet ont ete deja largement etudiees du point de vue descriptif
(Bornhaupt, 1867; Waldeyer, 1870; Gasser, 1874; Balfour & Sedgwick, 1879;
Gruenwald,
1941;
Kar, 1947; Abdel-Malek, 1950; Forsberg
&
Olivecrona, 1963),
l'analyse des relations entre cet organe et le canal de Wolff n'a fait Pobjet que de
travaux experimentaux peu nombreux. En 1937, Gruenwald, detruisant au
moyen de marques electrolytiques l'extremite distale du canal de
Wolff,
constate que toute interruption de ce dernier s'accompagne du blocage du canal
de Muller au meme niveau et en conclut que le canal de Wolff sert de 'rail
conducteur' a l'ebauche mullerienne en croissance. Les travaux ulterieurs
(Bishop-Calame, 1966; Didier, 1968a, b) confirment ces resultats et permettent
en outre de montrer (Didier, 19686) que le canal de Wolff induit la formation
de la plaque mullerienne, bande d'epithelium coelomique epaissi dont la dif-
ferenciation precede l'apparition du canal de Muller proprement dit et dont
l'extremite craniale invaginee est a
1'origine
du pavilion mullerien, Yostium tubae
abdominalis; ainsi que le soulignent Raynaud, Pieau & Raynaud (1970),
1
Adresse
de
Vauteur:
Laboratoire deBiologie animale, 1, Avenue Vercingetorix —63 —
Clermont-Ferrand, France.
8-2

116 E. DIDIER
j'emploie
ici le terme de 'plaque miillerienne' au sens large, pour designer non
seulement l'epithelium peritoneal epaissi de la region du futur ostium, mais
aussi celui qui se differencie dans le sens cranio-caudal le long du trajet du canal
de Wolff jusque dans la region cloacale, ces deux plages epitheliales etant en
continuity et ne presentant aucune difference du point de vue cytologique.
Dans cette perspective, il fallait done s'interroger sur le determinisme de la
formation de Vostium: la differentiation ostiale est-elle un phenomene auto-
nome, independant de l'uretere primaire?; ou bien l'ostium se forme-t-il sous
l'effet d'une influence inductrice emanant du canal de Wolff? C'est afin de
tenter de repondre a ces questions qu'a ete entrepris le present travail experi-
mental dont les resultats preliminaries ont fait l'objet d'une note prealable
(Didier, 1970).
Le principe de l'experience consiste a bloquer la progression du canal de
Wolff (voir Materiel et Methodes) en avant du territoire de l'ostium mullerien.
La localisation de la zone presomptive ostiale au niveau des somites 16 a 19 est
en effet connue depuis les observations histologiques d'Abdel-Malek (1950),
et sa mise en evidence par destruction elective chez l'embryon de 2,5 jours
d'incubation par les rayons X (Didier,
1968 a)
ou par experience de micro-
chirurgie (Didier, 1969). II
s'agit
done d'inhiber la penetration du blasteme de
l'uretere primaire en avant de la 16eme paire de somites et de verifier si l'agenesie
wolffienne et mesonephretique ainsi obtenue s'accompagne invariablement de
l'agenesie de Yostium mullerien.
MATERIEL ET METHODES
Dans ce but, 330 embryons de Poulet de race Hubbard ou provenant du
croisement entre Hubbard et Hybride Rhode Island
<£
x Wyandotte blanche ?,
ont ete operes unilateralement aux stades 8 a 13 de la table de developpement
de Hamburger & Hamilton (1951). Le blocage du canal de Wolff est assure par
un petit rectangle de membrane coquilliere d'oeuf de Poule non incube, introduit
le long de la bande somitique droite ou gauche, le cote oppose servant de cote
temoin (technique de Bishop-Calame, 1966). Les embryons operes sont sacrifies
entre le 6eme et le 15eme jour de l'incubation et etudies du point de vue macro-
scopique et histologique.RESULTATS
Sur les 116 embryons survivants etudies, 56 sont normaux (operation non
reussie), 60 presentent d'importantes anomalies du systeme uro-genital reparties
en cinq groupes principaux dans le Tableau 1.
1.
Formation du canal de Wolff et de Vostium
(a) Serie A
Chez 37 embryons operes entre la 12eme et la 19eme paires de somites,
Fintervention a ete realisee a une distance variable (evaluee en nombre de
somites presomptifs) de la derniere paire de somites differencies et se manifeste

Formation de r ostium
mullerien
117
par un blocage plus ou moins efficace du canal de Wolff au niveau de l'obstacle
(Tableau 2; Fig. 1A, B).
Ces resultats sont tout a fait comparables a ceux obtenus par Bishop-Calame
(1966);
cet auteur a en effet demontre la necessite d'operer a une distance d'au
moins trois somites en arriere du dernier somite differencie pour stopper
convenablement le canal de
Wolff.
Si Ton opere a une distance inferieure a trois
somites, l'obstacle sectionne le canal de Wolff dont la languette terminate
Tableau 1. Ensemble des resultats obtenus
Niveau de J'operation (nombre de somite) Nombre
, A v total
Resultat observe 11 12 13 14 15 16 17 18 19 de cas
(A) Canal de Wolff forme. — — 7567435 37
Ostium present
(B) Canal de Wolff forme. — — — —22 — — — 4
2 ouvertures ostiales
(C) Agenesie du canal de
Wolff.
1 6 1 — — 2 — — 1 11
Agenesie ostiale
(D) Agenesie du canal de
Wolff.
____ 2 — — — — 2
Ostium present
(E) Canal de Wolff forme. — — 2 1 1 — — 1 1 6
Ag6nesie ostiale
Tableau 2. Aspects de la morphogenese mesonephretique
chez les embryons de la serie A
Aspect de l'appareil urinaire
Destruction locale
Differenciation de du canal de
Wolff.
Interruption du tubules mesone- Differenciation
canal de Wolff phretiques en mesonephretique
au niveau de arriere de en arriere de
l'obstacle l'obstacle l'obstacle
Cote d'intervention... Droit Gauche Droit Gauche Droit Gauche
Distance d'intervention par rapport au dernier somite differencie
3 somites presomptifs 12 cas 5 cas 0 0 0 0
1 a 2 somites presomptifs 0 0 4 cas 5 cas 8 cas 3 cas
progresse en direction du cloaque, induisant sur son passage la differenciation
du blasteme mesonephrogene (Fig. 1B). Signalons toutefois que les elements
mesonephretiques ainsi differencies en arriere de l'obstacle sont frequemment
tres discrets, non visibles macroscopiquement (Fig.
1
A) et n'ont pu etre identi-
fies que sur les coupes histologiques ou ils se presentent sous la forme d'ilots de
trois ou quatre tubules.

118
E.
DIDIER

Formation de Vostium mullerien 119
Chez les 37 embryons de la serie A, Yostium mullerien a ete identifie dans tous
les cas le long du secteur du canal de Wolff forme. Tres rudimentaire chez les
embryons operes au niveau de la 13eme paire de somites, cet ostium se prolonge,
dans les autres cas, par un court canal de Muller de longueur variable bloque
dans sa croissance cranio-caudale soit par l'obstacle, soit par les deficiences de
l'appareil mesonephretique.
Les images de la Figure 2 illustrent la disposition relative des canaux de Wolff
et de Miiller en avant de l'obstacle et montrent l'agenesie wolffienne, mesone-
phretique et miillerienne en arriere du fragment de membrane coquilliere chez
un embryon opere au niveau du 14eme somite droit et sacrifie la 7eme jour de
l'incubation.
Tableau 3. Longueur moyenne
(JLL)
du segment du canal
de Muller edifie a cote du canal de Wolff
Age du sacrifice
5-6 jours
6-7 jours
7-12 jours
Somites
13 a 14
30
110
160
Niveau de l'operation
Somites
15 a 17
110
230
290
•\
Somites
18 a 19
1100
820
Remarque. Ainsi que je l'avais signale precedemment (Didier, 1969) les
formations urinaires et mulleriennes anterieures a l'obstacle se trouvent generale-
ment en position beaucoup plus craniale du cote opere que du cote temoin
(Fig. 1A). Cette disposition est probablement due au pincement des feuillets
embryonnaires par l'obstacle qui empeche le glissement en direction caudale de
l'ensemble du tractus uro-genital, et est vraisemblablement accentuee par
Fabsence de raccordement des voies uro-genitales au cloaque et l'agenesie des
ligaments musculaires qui maintiennent normalement en place les canaux de
Wolff et Muller.
Pour l'ensemble des embryons de cette serie, la longueur moyenne du secteur
mullerien forme a ete evaluee par denombrement
des
coupes transversales entre le
niveau de l'ouverture ostiale et l'extremite distale aveugle du canal de Muller
(Tableau 3).
Les chiffres montrent qu'en general, l'importance du secteur mullerien forme
FIGURE 1
(A et B) Appareil uro-genital de deux embryons de la serie A operes au niveau du
somite 16 et sacrifies a 9 jours. Presence d'ilots mesonephretiques (m) (non visibles
macroscopiquement chez l'embryon A) et de l'ostium mullerien (fleche).
(C
et D) Agenesies wolffienne, mesonephretique et ostiale totales chez
deux
embryons
de la serie C operes au niveau du somite
12
(sp,
septum pleuroperitoneal;
ov,
ovaire).
(C) Embryon opere a droite sacrifie a
10
jours.
(D) Embryon opere a gauche sacrifie
a
15
jours.
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%