Anatomie comparee

Anatomie comparée des différentes parties du cerveau chez les Vertébrés
1
Introduction
Pour évoluer dans leur environnement, les animaux ont besoin d’une structure centrale
qui reçoit l’information et élabore des réponses appropriées. Cette structure se décompose en
plusieurs parties : cerveau, tronc cérébral et moelle épinière. Nous ne prendrons pas en
compte l’évolution de cette dernière.
Les vertébrés ont en commun une chorde dorsale qui persiste à l’âge adulte, un rachis
ossifié pour protéger le système nerveux et un crâne ossifié pour mettre le cerveau à l’abri. On
sait aujourd’hui que ce dernier est à l’origine de l’élaboration des mouvements, de la
perception de l’environnement externe et interne ou encore du comportement social
indispensables à la survie de l’animal.
Pour répondre à toutes ces fonctions, il se décompose en 5 vésicules communes à tous
les vertébrés durant l’embryogenèse, mais dont l’évolution va varier selon les besoins de
l’adulte, notamment en fonction de son milieu (aquatique, aérien, terrestre).
Nous allons donc décrire le développement de chaque vésicule par l’étude de
l’anatomie de celles-ci, en fonction de la classification de l’individu et du milieu dans lequel il
évolue.

Anatomie comparée des différentes parties du cerveau chez les Vertébrés
2
I. Le Télencéphale : origine de l’olfaction et
facultés intellectuelles supérieures
Initialement vésicule de petite taille (télencéphale primaire), il se distend latéralement
en deux expansions latérales, les futurs hémisphères cérébraux, dont l'importance ira
grandissante des Cyclostomes aux Mammifères (télencéphale secondaire). (cf. figure 1)
Fig. 1 : Evolution de l’encéphale
Chez les Cyclostomes, il est rudimentaire et très peu développé par rapport aux autres
subdivisions de l'encéphale. Les hémisphères cérébraux se continuent vers l'avant par les
lobes olfactifs, très gros, d'où partent deux nerfs qui se rendent au sac olfactif impair.
Chez les Chondrichtyens et les Ostéichtyens, les bulbes olfactifs sont très gros et reliés
par des tractus. L’importance des bulbes olfactifs n’est cependant pas due au fait que leur sens
est plus développé que chez les hommes par exemple, mais au fait que les Poissons s’adaptent
à la faible teneur en molécules dissoutes. (cf. figure 2).
Fig. 2 : Encéphale de truite

Anatomie comparée des différentes parties du cerveau chez les Vertébrés
3
La structure de l'encéphale des Téléostéens est très différente de celle des autres
Vertébrés. Ce télencéphale est le résultat d'une « éversion » (=croissance vers l'extérieur) des
portions dorso-latérales du télencéphale primaire.
Chez les Reptiles, au niveau des hémisphères cérébraux, on constate l'apparition d'une
corticalisation des structures dorsales: il se différencie latéralement un paléocortex qui reste le
lieu des projections olfactives secondaires et médio-dorsalement un archéocortex qui est un
centre olfactif tertiaire. Le télencéphale devient de plus en plus volumineux et de plus en plus
complexe fonctionnellement, cela se traduit par une olfaction plus fine.
En revanche, on retrouve des lobes olfactifs très petits chez les Oiseaux qui ne
possèdent alors qu’une olfaction médiocre. Nous verrons plus loin que l’imperfection de ce
sens est compensée par le développement plus fin d’autres (notamment la vue). (cf. figure 3)
Fig. 3 : Encéphale de pigeon
Chez les Anamniotes et chez presque tous les Reptiles, la quasi-totalité du
télencéphale est au service de l’olfaction. A partir des monotrèmes, il apparaît de nouveaux
centres dans le cortex (pallium) ; à la partie ancienne ou archipallium s’en ajoute une nouvelle
appelée néopallium. La vue, l’ouïe et bien d’autres fonctions ont leurs centres réunis dans ce
néopallium dont le développement s’accompagne d’un accroissement des facultés
intellectuelles supérieures.
Les hémisphères prennent de l’ampleur dans de telles proportions qu’ils se replient sur
eux-mêmes, vers l’arrière, et recouvrent partiellement ou totalement (comme chez les
Primates) le reste de l’encéphale (cf. figure 4).

Anatomie comparée des différentes parties du cerveau chez les Vertébrés
4
Fig. 4 : Embryogenèse du néocortex
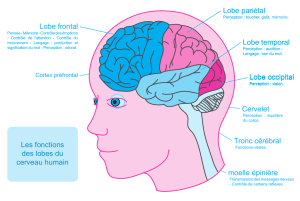
A l’achèvement de son développement, le néocortex se présente extérieurement
organisé en lobes où l’on reconnaît : un lobe pariétal pour la partie centrale, un lobe frontal
vers l’avant (particulièrement développé chez les Primates), un lobe temporal vers l’arrière et
tout à fait postérieurement un lobe occipital. Chez les Primates, les lobes frontal et temporal
recouvrent une portion du lobe pariétal qui constitue l’insula.
La surface du néocortex peut être lisse (lissencéphalie) ou se plisser en formant des
circonvolutions (gyrencéphalie). Dans les différents ordres on rencontre des formes des deux
types ; on accorde un niveau d’évolution plus marqué aux espèces gyrencéphales puisque
l’apparition de circonvolutions permet une augmentation de la surface du cortex dans un
espace restreint, et donc logiquement un meilleur traitement de l’information.
Le rat est lissencéphale et le mouton est gyrencéphale, cependant le premier est plus
évolué que le second. Par ailleurs, la baleine possède bien plus de circonvolutions que n’en
montre l’encéphale humain ! Cela est dû au fait que le cerveau est proportionnel à la taille de
la boîte crânienne.
L’embryogenèse retrace la phylogenèse : le développement des vésicules
télencéphaliques chez le fœtus humain retrace le chemin parcouru au cours de l’évolution.
Pour résumer, le télencéphale peut être considéré comme un cerveau olfactif chez les non
mammaliens. Des animaux aquatiques aux animaux aériens il passe d’un stade bulbe avec des
tractus à un stade bulbe avec des vrais nerfs crâniens (Ière paire des nerfs crâniens). La faible
teneur en molécules odorantes dans l’eau par rapport à l’air est à l’origine de ce phénomène.
Cependant on constate une exception chez les Oiseaux qui présentent une olfaction très
minime.
1
2
3
4
5
Insula
lobe temporal en
développement
sillon latéral
sillon central
sillon pariéto-
occipital
A
B
C
D
lobe frontal
lobe pariétal
lobe temporal
lobe occipital

Anatomie comparée des différentes parties du cerveau chez les Vertébrés
5
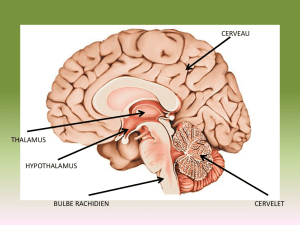
II. Le Diencéphale : régulateur de
l’homéostasie
Le diencéphale est à l’origine de trois structures. Il assure par l'intermédiaire du
thalamus dorsal la répartition des informations sensitives et par l’intermédiaire du thalamus
ventral un rôle moteur. Puis l'hypothalamus régule les fonctions physiologiques, il est
essentiel au maintien de l’homéostasie. Enfin, l’épithalamus forme la glande pinéale qui, par
ses sécrétions de mélatonine, régule les rythmes biologiques et la pigmentation de la peau.
Chez les Chondrichtyens et les Ostéichtyens, le thalamus est recouvert par une mince
toile choroïdienne (troisième ventricule) sur laquelle est couchée l'épiphyse, en arrière du
chiasma optique. Sa partie latérale et ventrale (thalamus) représente un centre régulateur des
grandes fonctions. Sa partie la plus ventrale (hypothalamus) est en relation anatomique et
fonctionnelle avec l'hypophyse.
Les poissons présentent un organe parapinéal réduit et un organe pinéal classique (ou
épiphyse) responsable de la détection de la lumière et de la photopériode. L’épiphyse sécrète
la mélatonine et est inhibée par la lumière, ce qui permet à l’animal d’être sensible aux
rythmes circadiens.
Chez les Reptiles, l’organe pinéal devient un « œil » alors qu’il est totalement absent
chez les Oiseaux puisqu’il n’est plus photosensible chez ces derniers. En effet c’est la rétine
qui va contrôler la sécrétion de mélatonine.
Chez les Anamniotes, le thalamus ventral est très développé et a un rôle moteur, il va
régresser chez les Amniotes au profit du cortex moteur plus élaboré.
Le thalamus dorsal quant à lui est un relais des voies sensorielles, il est très développé
chez les Mammifères et les primates. La masse du thalamus dorsal s'est accrue dans toutes les
directions du fait de son rôle de relais entre les étages postérieurs et le néocortex. Ce relais est
essentiellement assuré par les corps genouillés médians (C.G.M.) sur les voies de l'audition et
les corps genouillés latéraux (C.G.L.) sur les voies visuelles.
L'hypothalamus apparaît extérieurement plus simple que celui des poissons
(disparition des lobes inférieurs, du sac vasculaire...). En fait, son organisation est des plus
complexes chez les Mammifères car il est à l’origine des émotions, de l’apprentissage et de la
mémorisation.
On retiendra que le diencéphale est un centre d'intégration, de relais et de contrôle de
nombreux comportements assurant la survie de l'animal (faim, soif, comportement sexuel);
qu'il assure au plus haut niveau la régulation des processus homéostatiques tels que la
pression artérielle, la régulation thermique (homéothermie), l'équilibre de l'eau et des sels
minéraux. Enfin, il apparaît comme un centre d'intégration supérieur du système
neurovégétatif et du système endocrinien par le biais de l'hypophyse.
Le diencéphale est présent chez toutes les classes car il a un rôle essentiel dans le
contrôle des fonctions viscérales et endocrines. Chez certains groupes, on retrouve d’autres
fonctions comme la régulation du cycle nycthémère, la capacité à ressentir des émotions.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%