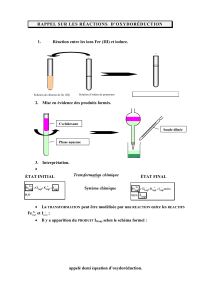
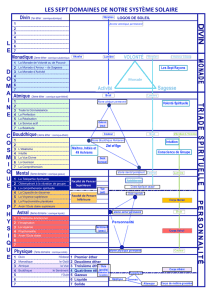
Un rôle physiologique des plasmalogènes : la - iPubli

SYNTHÈSE
---,

b
95
1 �
1

1

0
H0 -0
XH
� )2 o
HO
0
0
2.
à
1
J+)

0
!
[
"
CH
TSH
»
�
3.
 6
6
1
/
6
100%