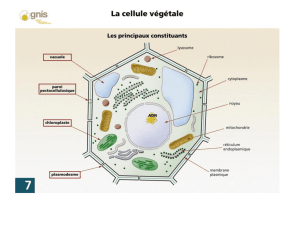
La vacuole

1
LA VACUOLE
I/Introduction.
La vacuole (vacuus=vide) :
- Occupe la majorité du volume cellulaire (80%).
- Contient de très nombreuses molécules et ions dissouts.
- Entretien de constants échanges avec le cytoplasme rôle régulation.
- Toujours en turgescence.
Entre cytoplasme et vacuole, on trouvera une bicouche de lipide, le tonoplaste, qui va réguler les
échanges de la vacuole avec le cytoplasme.
1) La pression osmotique.
Les cellules stomatiques permettent les échanges gazeux par l’ouverture de l’ostiole qui se fait par
la pression vacuolaire :
- Ouverture = turgescence.
- Fermeture=plasmolyse.
La pression osmotique est:
- Proportionnelle au nombre de molécules et d’ions dissouts.
- Toujours importante, dépasse la pression atmosphérique (2 et 20 atm).
Le gène vacuoleless régule le développement vacuolaire.
Les lois de l’osmose : les mouvements d’eau se font toujours vers le milieu le plus concentré
pompage de l’eau par la plante à 100 atm (en milieu aride).
II/Le contenu vacuolaire.
La vacuole contient principalement des ions et des molécules organiques.
1) Les produits du métabolisme primaire.
Les acides carboxyliques sont les molécules les plus importantes de la vacuole dont le pH se
maintient à 5.5 (100 fois plus de H+ que dans le cytosol).

2
a) Les acides carboxyliques :
Les différents acides présents :
- L’acide citrique (triacide) : est stocké dans la vacuole en absence de demande cellulaire.
- L’acide malique (diacide) : compartimente le CO2 dans la vacuole puis le libère pour la
photosynthèse (rôle majeur dans les plantes grasses). Cycle de Calvin : Malate pyruvate +CO2.
- L’acide oxalique (biacide) :
Composé toxique (rôle de défense).
Stocké sous forme double réduite (COO2-), associé à des Ca2+.
Acide glycolique acide glyoxyliqueacide formiqueoxalate.
b) Les glucides :
Le saccharose :
- Présent en grande quantité dans la vacuole.
- Il est issu de la photosynthèse.
- Stocké à court terme en périodes de fortes activités photosynthétiques.
- Rôle de transport des glucides dans la sève (ne peut être réduit).
Réserves glucidiques dans les tubercules pour redémarrer le métabolisme.
Il y a aussi stockage d’amidon mais celui-ci est stocké dans les plastes et jamais dans la vacuole.
c) Les acides aminés :
Il y a peu de protéine au sein de la vacuole (sauf exception), elle ne contient que de 5% du volume
protéique de la cellule.
Ce sont principalement des enzymes hydrolytiques capables de dégrader des molécules (protéases
et estérases).
Les légumineuses permettent de maintenir le nombre d’AA dans le cytoplasme.
La rétroinhibition contrôle la synthèse des AA par les précurseurs, les AA non utilisés sont stockés
dans la vacuole et ne permettent pas la rétroinhibition : AAprécurseurs.
Régulation du nombre d’AA dans la cellule.
2) Les composés du métabolisme secondaire.
La solubilisation des molécules pigmentaires :
- Se fait par attache des glucides aux groupements OH des pigments.
- Libération des H+ par la réaction d’estérification.
- Augmente le pH vacuolaire.
Le pH est donc un déterminant de la couleur de la plante.
Les molécules du métabolisme secondaire:
- Les anthocyanes sont responsables de la couleur des fleurs (=pigments flavonoïdes).
- Les alcaloïdes sont capables de bloquer les mitoses (nicotine).
- Les tannins et les alcaloïdes sont des dénaturants protéiques (tannage du cuir).
Les vacuoles stockent les pigments car le signal de couleur attire les insectes pollinisateurs.

3
3) Les vacuoles des graines.
Leurs vacuoles possèdent un grand nombre de protéines qui sont dégradés en AA et permettent le
développement de la plante.
Cette maturation est accompagnée d’une déshydratation qui fragmente la vacuole (≠germination).
La synthèse protéique :
- Les protéines sont synthétisées avec une séquence additionnelle : peptide signal (coté Nterm).
- Reconnue par la membrane du REG et y est intégrée.
- Coupée et dégradée par la signal peptidase.
Adressage des protéines à la vacuole :
- Amenées par flux vésiculaire exocytaire.
- Possèdent des séquences d’adressage SP + NTPP ou CTPP (sinon adressage à la paroi).
Les protéines de la vacuole sont codées par un gène identifié.
Les graines stockent les enzymes et leurs substrats dans la vacuole pour effectuer la dégradation
des protéines.
La maturation qui entraine la fragmentation de la vacuole permet le stockage des enzymes et des
substrats dans différentes petites vacuoles.
Les vacuoles PSV stockent les protéines.
Les vacuoles CCV (lyriques) stockent les vésicules de clathrine.
Les corps protéiques peuvent être formés par autophagie de la vacuole (concerne surtout les
protéines de réserves=prolamines).
III/Le tonoplaste.
Le tonoplaste est la membrane vacuolaire (8nm) qui s’agrandit par voie vésiculaire (pas
d’autosynthèse), et permet les échanges vacuole-cytoplasme.
1) Les activités de transports.
Le tonoplaste possède les mêmes propriétés que la membrane plasmique :
- Imperméable aux molécules hydrophiles (petites exceptées).
- Perméables aux molécules hydrophobes.
- Transport effectué par les protéines intrinsèques.
Présente des différences de charges selon les feuillets.
Transport passif par gradient électrochimique : les molécules suivent les différences de charges.
Transport actif se fait uniquement par apport d’énergie (sens inverse des charges).
Transport multimoléculaire en même temps et dans le même sens : symport.
Transport multimoléculaire en même temps mais de sens opposé : antiport.
Les dispositifs de régulation d’ouverture des canaux sont:
- Voltage dépendant.
- Mécanosensible.
Il y a des récepteurs spécifiques aux molécules qui se présentent à l’entrée des canaux.

4
a) Les ATPases à protons et pyrophosphatases :
C’est un complexe de protéine intrinsèque qui permet le transport de proton.
Le gradient électrochimique va de la vacuole au cytoplasme.
La pompe à H+ contient deux parties:
- Une partie fixe : stator.
- Une partie mobile : rotor.
Présente un transport actif de proton du cytoplasme à la vacuole donc nécessite de l’énergie.
Les pyrophosphatases sont des systèmes de protéine intrinsèque qui tirent leur énergie de
l’hydrolyse du pyrophosphate.
Ces systèmes entrainent la création d’un gradient de protons.
b) Les transports dépendant du gradient de proton :
Le transport du Ca2+:
- Transport actif et antiport.
- Transport primaire des H+ par la pompe ATPase.
- Permet l’entrée du Ca2+ par gradient de protons (augmentation [Ca2+] en présence d’ATP).
La vacuole est un réservoir de calcium pour la cellule : transport régulier.
Les ionophores à H+ sont des molécules chimiques capables de permettre le passage d’un ion et de
perméabiliser la membrane aux protons :
- Plus de gradient de H+.
- Entrée du Ca2+ stoppé.
Pas de transport antiport possible.
Le transport du malate se fait sous forme doublement réduite par un récepteur spécifique pour
suivre le gradient électrochimique et se protonise ensuite.
c) Les transporteurs ABC :
Les ATP Binding Casette permettent l’entrée de molécules du métabolisme secondaire par
hydrolyse d’ATP directement dans la vacuole.
Ces molécules transportées sont souvent couplées au glutathion (Glu-Cys-Gly).
La glutathion-transférase (GST) permet la conjugaison de la molécule au glutathion.
Dans le cas de l’anthocyane il n’y a pas de transport bidirectionnel.

5
d) Le transport de l’eau :
Le transport de l’eau se fait par des aquaporines qui sont des canaux d’eau intrinsèques à la
membrane, ils sont extrêmement sélectifs et ne laissent pas passer les ions.
Les molécules d’eau passent une à une à l’intérieur de ce pore.
Stabilise la pression osmotique sans dérégler le rapport ionique.
2) La formation de la vacuole.
La vacuole se forme à partir du REG qui créé plusieurs petites vacuoles lysosomales qui fusionnent
Les méristèmes sont des cellules indifférenciées qui donnent naissance à toutes les cellules
végétales : ce sont des cellules totipotentes.
3) Les rôles de la vacuole.
La croissance cellulaire : c’est le compartiment qui croit le plus dans la cellule.
Maintient l’homéostasie du cytoplasme :
Digestion.
Stockage.
Echanges sélectifs.
Défense de la cellule : les composés cyanogénétiques libèrent du cyanure qui se fixe sur les
cytochromes des mitochondries et empêchent la respiration cellulaire.
1
/
5
100%