Chapitre 7 : Acoustique physiologique

Chapitre 7 : Acoustique
physiologique
1 Introduction à l’acoustique physiologique et à la psychoacoustique
Puisque c’est par l’intermédiaire de l’oreille que le son est perçu, et que la sensation auditive
est interprétée par le cerveau, l’étude des sons ne relève pas uniquement de la physique,
mais aussi de la physiologie de l’oreille et de la neurophysiologie (acoustique physiologique)
et de la psychologie (psychoacoustique).
Ainsi, de nombreuses contingences physiologiques et psychologiques limitent la perception
du phénomène auditif, par exemple :
Un son intense peut en cacher un autre (effet de masque).
La perception d’un son bref dépend de la durée du stimulus.
La perception auditive varie d’un sujet à l’autre, en fonction de l’âge, du sexe, et de
l’état médical du sujet.
Nous disons que nous entendons un son lorsque des vibrations de l'air ambiant, atteignant
notre tympan, le mettent en mouvement dans des conditions d'amplitude et de fréquence
telles que cette stimulation mécanique, qui est transmise par l'oreille moyenne à l'oreille
interne, y provoque un phénomène bio-électrique.
Commence alors le traitement de l'information contenue dans ce phénomène, traitement
qui se poursuit à travers différents relais jusqu'au cortex cérébral et dont le résultat sera la
perception du son.

L’oreille a une sensibilité très importante : un déplacement du tympan de 10-9 cm
(inférieur au diamètre de la plus petite molécule de l’air, celle de H2) provoque un effet
sensible.
L’oreille est sensible à des intensités acoustiques très différentes (le rapport extrême des
intensités audibles équivaut à 1013, voire 1014).
L’oreille n’est pas sensible à toutes les fréquences acoustiques (seuils absolus de
fréquence).
Le spectre des fréquences audibles est très étendu ; il va de 16 à 16 000 Hz, et couvre
donc 10 octaves.
L’oreille ne différencie pas deux fréquences trop proches l’une de l’autre (seuil différentiel
de fréquence).
Nous abordons dans ce chapitre la partie acoustique physiologique. La psychoacoustique
fera l’objet d’un autre chapitre.

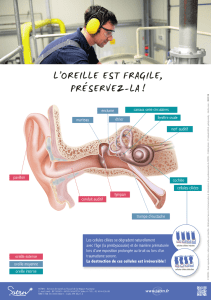
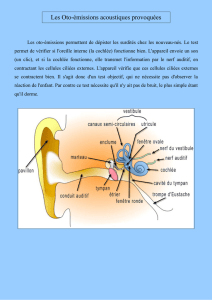
2 Anatomie et physiologie de l’oreille
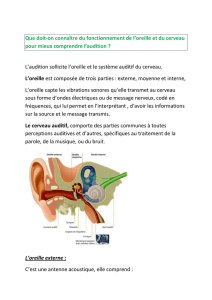
On distingue trois parties consécutives dans l'appareil auditif :
l’oreille externe (formée du pavillon et du conduit auditif),
l’oreille moyenne (limitée par le tympan et les fenêtres de la cochlée),
l’oreille interne (renfermant les organes de l’audition, situées dans la cochlée, et les
organes du maintien de l’équilibre, les canaux semi-circulaires).


 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
1
/
196
100%