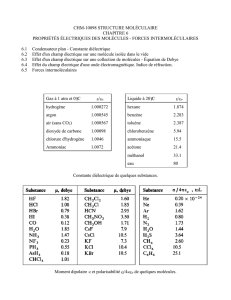
ACTIVATION ET POLARISATION DES LYMPHOCYTES T

UE 5 Immunologie -Cours 9
Pr Toubert
Jeudi 8 Décembre à 8h30
Ronéotypeuse : Laura Mattei
Ronéolecteur : Antoine Allouche
ACTIVATION ET POLARISATION DES
LYMPHOCYTES T

SOMMAIRE
I. Introduction
II. Phase d’initiation
III. Expansion clonale
IV. Phase effectrice.
1) production de cytokines
2) les lymphocytes T helper
2) la fonction cyto-toxique

I Introduction :
La différence entre immunité innée et immunité acquise?
L'immunité innée c'est l'ensemble des réponses qui se produisent d'emblée dans la
première reconnaissance d'un pathogène, elle reconnaît des structures conservées entre
différents pathogènes (PN, monocytes, macrophages, NK).
L'iimmunité acquise ou adaptative c'est la reconnaissance préalable du pathogène après
sa présentation, qu'il s'agisse de peptides présentés par les molécules du CMH ou
d'épitopes antigéniques (sur les protéines bactériennes par exemple), qui sont reconnus
par des récepteurs spécifiques : des Ig (LB) et TCR (LT)
L’immunité acquise se met en route en 3 phases :
- l’initiation par la coopération entre une cellule présentatrice d’antigène et un lymphocyte
T.
- l’expansion clonale et coopération cellulaire
coopération entre Ly. Th et Ly. B
coopération entre Th et autres Ly. T
- enfin, la phase effectrice avec :
les différents lymphocytes helper: Th1, Th2 et cytokines
les cellules cytotoxiques (CTL ou CD8)
II. Phase d’initiation
L’initiation de la réponse immune a lieu dans les organes lymphoïdes secondaires :
ganglions, rate, système lymphoïde muqueux (MALT). La réponse T est initiée par la
cellule présentatrice d’Ag (CPA) activée.
C’est une cellule qui présente sur sa membrane, de façon constitutive, des molécules du
complexe majeur d’histocompatibilité de classe 2 et va permettre la maturation des LT
naïfs (de phénotype CD4 et CD8).
On distingue 3 types de cellules présentatrices d’antigène :
- les cellules dendritiques. Ce sont les cellules les plus importantes car elles permettent
l’activation des LT naïfs. Elles rencontrent l’antigène et le phagocytent pour présenter un
peptide antigénique au LT qui est alors naïf.
- les LB.
- les macrophages.
Le lymphocyte passe ainsi de LT naïf à LT effecteur. On a différents types de LT effecteur :
• T helper (CD4)
TH1 : immunité cellulaire
TH2 : production d ’anticorps, coopération T:B
• T cytotoxique (CD8)

Les CPA professionnelles ont différentes caractéristiques, elles doivent être capables de :
•synthétiser des molécules du CMH II
•capturer un Ag
•dégrader l’Ag en fragments peptidiques
•charger le peptide sur les molécules du CMH II
•exprimer les molécules de co-stimulation.
Rappel sur les molécules du CMH:
Les molécules de classe I et de classe II se ressemblent beaucoup, elles ont une
organisation globalement similaire.
Cellules de classe I : elles sont ubiquitaires, présentes sur la plupart des cellules
nucléées, mais néanmoins très peu exprimées dans le SNC, et sont constituées d'une
chaine lourde plus la microglobuline
Cellules de classe II : deux chaines et reliées de façon non covalente. Elles s'occupent
des Ag exogènes présents dans l'environnement de l'Ag qui sont captés par des
mécanismes d'endocytose.
La biosynthèse de la molécule de classe II est faite par le RE puis elle va être diriger dans
des endosomes primitifs. Le peptide est déplacé par des molécules pour aboutir à une
molécule de classe II fixant le peptide qui sera présenté à la surface.
La première étape de l’initiation est la rencontre de l’antigène avec la cellule dendritique
immature en périphérie.
C’est une cellule très performante dans la phagocytose de l’antigène, mais très peu pour
la présentation de l’antigène et enfin elle exprime très peu sur sa surface des molécules
de co-stimulation . Donc notre cellule capte l’antigène, puis exprime ensuite à sa surface
les CMH de classe I et II et augmente la présence membranaire des molécules de co-
stimulation.
NB : cette maturation des cellules dendritiques ne se fait pas à un point donné mais est
progressive pendant leur migration vers les organes lymphoïdes secondaires.

schéma où tu es sensé voir la maturation des cellules dendritiques
Donc la CD immature (tissu) a une capacité de capture importante mais une capacité
moindre d'expression des CMH et des molécules de co-stimulation.
A l'inverse la CD mature (ganglion), elle, perd sa capacité de capture et va exprimer en
plus grandes quantités les molécules de CMH I et II et de co-stimulation.
La deuxième étape est le contact de la cellule dendritique mature et d’un LT naïf dans le
ganglion. En effet, il y a migration des CPA qui ont capté l'Ag qui vont vers le ganglion
pour rencontrer le LT et l'activer.
A savoir que la cellule dendritique immature, qui acquière sa maturation en migrant par
voie lymphatique, va exprimer des molécules d’adhésion pour amorcer son entrée dans la
zone para-corticale du ganglion. Le LT entre dans le ganglion par un mécanisme de
homing et là s’effectue des contacts aléatoires entre LT naïfs et cellules dendritiques
matures ou activées.
NB : bien se rappeler que la cellule dendritique est activée en périphérie et qu’elle mûrit en
migrant, mais une fois dans le ganglion elle est mature (capacité réduite de phagocytose,
une grande capacité de présentation de peptide antigénique et une expression augmentée
de molécules de co-stimulation.) Donc on a deux possibilités : soit la cellule dendritique
rencontre le LT naïf spécifique (= TCR spécifique) de l'Ag qu'elle porte et le LT va devenir
effecteur à plus courte durée de vie; ou alors le LT ne l’est pas et celui ci sort du ganglion.
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%