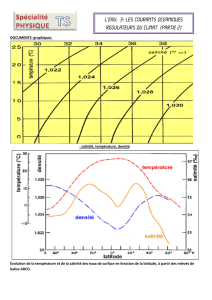
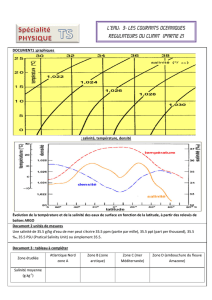
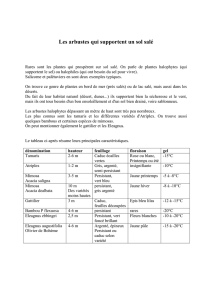
tableau 1 - Università di Sassari

UNIVERSITÀ DEGLI STUDI DI SASSARI
Facoltà di Agraria
Dottorato di Ricerca in Agrometeorologia ed Ecofisiologia
dei Sistemi Agrari e Forestali
XX ciclo
ADAPTABILITE DES ESPECES DU GENRE ATRIPLEX
AUX CONDITIONS DE SALINITE ET D'ARIDITE
Coordinatore:
Chiar.mo Prof. Pietro Deidda
Dottorando: Dott.ssa Oumelkheir Belkheiri
Docente guida:
Prof. Maurizio Mulas
Anno Accademico 2007/2008

B
Be
el
lk
kh
he
ei
ir
ri
i
O
Ou
um
me
el
lk
kh
he
ei
ir
r,
,
Adaptabilité des espèces du genre Atriplex aux conditions de salinité et d’aridité, Tesi di
D
Do
ot
tt
to
or
ra
at
to
o
i
in
n
A
Ag
gr
ro
om
me
et
te
eo
or
ro
ol
lo
og
gi
ia
a
e
ed
d
E
Ec
co
of
fi
is
si
io
ol
lo
og
gi
ia
a
d
de
ei
i
S
Si
is
st
te
em
mi
i
A
Ag
gr
ra
ar
ri
i
e
e
F
Fo
or
re
es
st
ta
al
li
i,
,
U
Un
ni
iv
ve
er
rs
si
it
tà
à
d
di
i
S
Sa
as
ss
sa
ar
ri
i

A
A
l
la
a
m
mé
ém
mo
oi
ir
re
e
d
de
e
m
mo
on
n
p
pè
èr
re
e
A
A
t
to
ou
us
s
c
ce
eu
ux
x
q
qu
ui
i
m
me
e
s
so
on
nt
t
c
ch
he
er
rs
s
B
Be
el
lk
kh
he
ei
ir
ri
i
O
Ou
um
me
el
lk
kh
he
ei
ir
r,
,
Adaptabilité des espèces du genre Atriplex aux conditions de salinité et d’aridité, Tesi di
D
Do
ot
tt
to
or
ra
at
to
o
i
in
n
A
Ag
gr
ro
om
me
et
te
eo
or
ro
ol
lo
og
gi
ia
a
e
ed
d
E
Ec
co
of
fi
is
si
io
ol
lo
og
gi
ia
a
d
de
ei
i
S
Si
is
st
te
em
mi
i
A
Ag
gr
ra
ar
ri
i
e
e
F
Fo
or
re
es
st
ta
al
li
i,
,
U
Un
ni
iv
ve
er
rs
si
it
tà
à
d
di
i
S
Sa
as
ss
sa
ar
ri
i

R
RE
EM
ME
ER
RC
CI
IE
EM
ME
EN
NT
TS
S
Ce travail a été réalisé à la station expérimentale du département d’Economie et
des Systèmes Arborées DESA sise à Fenosu, Oristano et aux laboratoires du DESA,
d’Agropedologie et de Zootechnie de la faculté d’agriculture de l’université de Sassari,
sous la direction du Pr. Maurizio Mulas, Professeur au DESA, Section Ecosystèmes
Agraires et Arborés. Qu’il trouve ici l’expression de ma profonde reconnaissance pour
m’avoir fait bénéficier de ses connaissances et de sa grande expérience ainsi que pour la
confiance qu’il a bien voulu m’accorder en me proposant ce sujet.
Ce travail n’aurait pas vu le jour sans le soutien financier du ministère des
affaires étrangères Italien sollicité par le Pr. Giuseppe Enne que je tiens spécialement à
remercier pour m’avoir accueillie au sein du Nucleo Ricerca alla deserificazione NRD,
pour l’intérêt qu’il a manifesté envers mon travail et pour son soutien moral tout au long
de mon parcours. Merci pour tout.
Je tiens à remercier vivement Pr. Pietro Deidda et Pr. Innocenza Chessa pour
avoir accepté d’examiner ce travail et pour leur conseils très constructifs. Merci encore.
Mes vifs remerciements vont aussi à Pr. Antonello Cannas, directeur du
département de Zootechnie pour son appui scientifique, Mr. Roberto Rubato pour
m’avoir encadrée et orientée et à tout le personnel du département de zootechnie, pour
leur aide et soutien durant mon bref séjour au laboratoire d’analyses des aliments.
Je tiens à exprimer toute ma gratitude à Mario Deroma du département
d’agropédologie pour son aide précieuse lors de la réalisation du travail expérimental.
Je n’oublie certainement pas de remercier Mr.Giovanni Ligios et Mr Filippo
Virdis du laboratoire de chimie du DESA pour leur aide et leur patience durant mon
travail expérimental.
Un grand merci à Giovanna Seddaiu et Luigi Ledda pour leur aide précieuse
malgré leur emploi du temps toujours chargé.
B
Be
el
lk
kh
he
ei
ir
ri
i
O
Ou
um
me
el
lk
kh
he
ei
ir
r,
,
Adaptabilité des espèces du genre Atriplex aux conditions de salinité et d’aridité, Tesi di
D
Do
ot
tt
to
or
ra
at
to
o
i
in
n
A
Ag
gr
ro
om
me
et
te
eo
or
ro
ol
lo
og
gi
ia
a
e
ed
d
E
Ec
co
of
fi
is
si
io
ol
lo
og
gi
ia
a
d
de
ei
i
S
Si
is
st
te
em
mi
i
A
Ag
gr
ra
ar
ri
i
e
e
F
Fo
or
re
es
st
ta
al
li
i,
,
U
Un
ni
iv
ve
er
rs
si
it
tà
à
d
di
i
S
Sa
as
ss
sa
ar
ri
i

Un grand merci à tous mes amis, thésards, travailleurs et stagiaires de la faculté
d’agriculture et tout particulièrement à Rita Melis pour son soutien moral et logistique,
Michele Salis pour sa sympathie et sa gentillesse, Serena, Maurizio, Giovanni et Mauro
pour m’avoir fait sentir que j’étais parmi les miens. Merci encore.
Je prie tout le personnel de la station expérimentale d’Oristano de trouver ici
l’expression de ma profonde gratitude pour leur participation effective à la réalisation de
ce travail.
Je remercie tout le staff du NRD et particulièrement mes amis Helmi Saidi pour
son appui technique, Roberta Lobina, Nawal El Azhari, Davide Bellavite, Marcello
Lubino pour leur aide, leur soutien de tous les instants et leurs encouragements.
Que tous ceux qui ont contribué de prés ou de loin à la réalisation de ce travail et
que je n’ai pas cités me pardonnent et trouvent exprimés ici mes vifs remerciements.
B
Be
el
lk
kh
he
ei
ir
ri
i
O
Ou
um
me
el
lk
kh
he
ei
ir
r,
,
Adaptabilité des espèces du genre Atriplex aux conditions de salinité et d’aridité, Tesi di
D
Do
ot
tt
to
or
ra
at
to
o
i
in
n
A
Ag
gr
ro
om
me
et
te
eo
or
ro
ol
lo
og
gi
ia
a
e
ed
d
E
Ec
co
of
fi
is
si
io
ol
lo
og
gi
ia
a
d
de
ei
i
S
Si
is
st
te
em
mi
i
A
Ag
gr
ra
ar
ri
i
e
e
F
Fo
or
re
es
st
ta
al
li
i,
,
U
Un
ni
iv
ve
er
rs
si
it
tà
à
d
di
i
S
Sa
as
ss
sa
ar
ri
i
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
1
/
182
100%