COURS D`IMMUNOLOGIE GENERALE (Dr. Adom) Année

1
COURS D’IMMUNOLOGIE GENERALE
(Dr. Adom)
Année Universitaire 2013-2014
CHAPITRE I : GENERALITES
A – QUELQUES DEFINITIONS ET RAPPELS HISTORIQUES.
La vie de chaque organisme vivant est constamment menacée par d’autres/ C’est la nature
même du monde vivant. En réponse à cette agression constante, chaque espèce vivante a
perfectionné un mécanisme de protection allant du camouflage colorié de certains poissons,
au développement musculaire pour la course.
Dans leur combat continu contre les micro-organismes, les vertébrés ont élaborés un
ensemble de mesures de protection appelés « Système Immunitaire ».
La clé de voûte du système immunitaire est le processus de reconnaissance : le système
immunitaire doit être capable de faire la distinction entre les constituants naturels du corps
(soi) et un corps étranger (non soi). Toute défaillance dans ce processus de reconnaissance
peut conduire à des troubles graves connues sous le terme de « maladies auto-immunes »
dont les formes les plus sévères sont fatales.
L’Immunologie est la discipline des Sciences qui s’occupe de l’étude de l’immunité.
L’Immunité, c’est la capacité à résister, à limiter le développement, à se débarrasser d'une
maladie après avoir été infecté par un agent étranger (bactéries, champignons, virus,
parasites). C’est l’ensemble des facteurs humoraux et cellulaires qui protègent l'organisme
contre une agression infectieuse ou toxique. Le terme d’immunité est à l'origine juridique. Il
vient du latin in munis et désigne un individu exempté de charges. En effet, au Moyen-âge,
on savait que le sujet guéri d'une maladie contagieuse au cours d'une épidémie, ne
contractait plus cette même maladie au cours des épidémies ultérieures : cet individu
pouvait dès lors être mobilisé pour soigner les malades, en compensation de quoi il était
exempté de charges (d’impôts). Un animal qui est immunisé contre un agent pathogène
spécifique est « libéré » de toute infection par ce micro-organisme.
L’immunité peut être naturelle ou acquise.
L’Immunité Naturelle ou Innée est la capacité, pour un individu, de posséder une résistance
naturelle à un grand nombre d'agents infectieux. Cette forme d’immunité est possédée dès
la naissance du fait de caractères constitutionnels multiples sous contrôle génétique.
L’Immunité Acquise, comme son nom l’indique, est acquise au cours de la vie. Elle est
spécifique d'un seul type de microorganismes.
Le premier écrit se rapportant à l'acquisition d'une résistance vis-à-vis d'une maladie
infectieuse paraît être celui de Thucydides (historien Grec, 465-395 avant J.C, qui s'intéresse
aux faits sociaux et à leurs causes). Il concerne la peste qui sévissait à Athènes. Le premier
écrit relatant la production intentionnelle d'une immunité spécifique, est celui de Voltaire,
qui dans une lettre de 1733, rapporte que dès le XIème siècle, les chinois pour se protéger
contre la variole, s'inoculaient la maladie en inoculant de la poudre de croûtes desséchées
de varioleux. Cette pratique, en général, déterminait une forme atténuée de l'affection, mais
il y avait aussi le risque de faire une forme grave.
Ce procédé nommé variolisation ou variolation a été introduit en Europe en 1721.

2
En 1798, Jenner propose un procédé moins dangereux, l'inoculation d'une maladie bénigne
pour l'homme, la vaccine ou variole des bovidés pour le protéger contre une affection
humaine grave, la variole.
Par la suite, Pasteur pour une Pasteurellose mortelle des Gallinacées (le choléra des poules),
montre que ces animaux peuvent devenir résistants à cette maladie en leur injectant une
vieille culture du germe dans laquelle l'agent pathogène a perdu une partie de sa virulence.
Pasteur en hommage à Jenner, nomme cette méthode de protection, utilisant l'injection de
germes vivants peu ou pas pathogènes, la vaccination (pour rappeler que Jenner avait fait
ses premiers essais avec la vaccine).
La vaccination jennerienne utilise des germes vivants peu ou pas pathogènes voisins du
germe responsable de l'affection contre laquelle on veut lutter. La vaccination pasteurienne
fait appel à l'agent pathogène à l'origine de la maladie mais auquel, sans le tuer, on a fait
perdre ou diminuer artificiellement son pouvoir pathogène. Le premier vaccin de ce type
chez l'homme est le vaccin antirabique.
Plus tard, en 1886, Salmon et Smith constatent que le bacille tué du choléra des poules est
aussi efficace dans la protection que le bacille atténué de Pasteur. Ainsi, naissait la
vaccination par germes tués par opposition à la vaccination par germes vivants de Pasteur
et Jenner. Par la suite, certains constituants du germe, comme les toxines sous forme non
toxique (anatoxines), ont été utilisés comme vaccin. Enfin, actuellement on propose
d'injecter au sujet à vacciner l'ADN du germe associé à un vecteur, pour faire produire par
les propres cellules de ce sujet les constituants du germe capables de provoquer une
immunité.
B – LES ACTEURS IMPLIQUES DANS LA REACTION IMMUNITAIRE.
Les acteurs impliqués dans la réaction immunitaires sont :
les organes,
les lymphocytes,
les anticorps,
les antigènes,
les récepteurs pour l’antigène,
les cellules présentatrices d’antigènes,
les molécules du Complexe Majeur d’Histocompatibilité (CMH),
les marqueurs CD (Cluster of Differentiation),
les ligands ;
les cytokines.
Tous ces acteurs vont êtres mobilisés pour une réponse innée ou spécifique (quoique non
exclusives, l’une de l’autre). Nous les aborderons au fur et à mesure du développement du
cours. Pour l’instant, nous présentons dans leurs grandes lignes, quelques uns parmi ces
acteurs.
B-1 Les organes de l’immunité
La moëlle osseuse,
le thymus,
la rate,
les ganglions lymphatiques et
le tissu lymphoïde

3
sont le lieu de production et de différenciation des cellules de l'immunité. On peut les ranger
en deux catégories : les organes centraux et les organes périphériques.
1) Les organes centraux
Le thymus peuplé de lymphocytes (appelés pour cette raison lymphocytes T) dont la
majorité reste dans le thymus et une partie (5 %) circule dans le sang et la lymphe.
La moëlle osseuse (dont l’équivalent chez les oiseaux est la Bourse de Fabricius) est le
lieu de production de toutes les cellules de l’immunité et des cellules hématopoïétiques.
Les lymphocytes qui la peuplent sont appelés lymphocytes B (en anglais, moëlle osseuse
= bone-marrow).
2) Les organes périphériques
Les organes lymphoïdes périphériques sont :
les ganglions lymphatiques,
la rate,
les formations lymphoïdes annexées au tube digestif (GALT : gastro-enteric associated
lymphoid tissues) ou au système respiratoire (BALT : bronchus associated lymphoid
tissues).
Les lymphocytes T et B les habitent dans des territoires qui leur sont réservés. On y trouve
également des macrophages. Ces organes lymphoïdes périphériques sont le siège de la
réaction immunitaire.
Trois types de facteurs interviennent pour protéger le sujet immunisé contre la maladie : les
anticorps (Ac), le complément (C) et différentes cellules.
B-2 Les anticorps (Ac)
En 1888, l'Ac est le premier facteur de l'immunité découvert. Il l'est par Roux et Yersin : ces
auteurs montrent que l'injection de l'exotoxine du bacille diphtérique entraîne l'apparition
dans la circulation sanguine d'une substance antitoxique spécifique. On parle alors d’une
Intestin grêle

4
immunité antitoxique. Le transfert du sérum de l'animal immunisé à un animal sain rend ce
dernier insensible à l'action toxique de la toxine. La même constatation est faite pour la
toxine tétanique par Behring et Kitasato (1890). Dans ces deux cas, on a transféré
passivement l'immunité. Le substratum de cette immunité antitoxique est un facteur sérique
synthétisé par l'organisme : l'anticorps (Ac). Cet Ac est spécifique de la molécule qui a induit
sa production et cette molécule porte le nom d'antigène (Ag) et on dit qu'elle est
antigénique ou immunogène.
A côté de cette immunité antitoxique, il existe l’immunité antibactérienne. Charrin et Roger,
en 1889, démontrent que si on cultive les Pseudomonas aeruginosa en présence d'un sérum
provenant d'un animal inoculé avec cette bactérie, une autre variété d'Ac agglutinent les
Pseudomonas aeruginosa. En 1896, Widal utilise cette agglutination pour faire le diagnostic
des Salmonelloses : c’est ainsi que nait le sérodiagnostic de Widal et Félix. En 1897, Krauss
met en évidence des Ac responsables du phénomène de précipitation. Celui-ci se produit si
le sérum d'un sujet immunisé est mélangé à un filtrat de culture du germe qui a servi à
l'immunisation. Donc, existent des Ac neutralisants, bactériolytiques, agglutinants et
précipitants, qui se définissent par des propriétés biologiques ou physiques différentes. Les
auteurs de cette époque ne savaient pas si à chacune de ces propriétés correspondait un
type particulier d'Ac ou si chaque Ac, comme c'est en fait le cas, possédait toutes ou
certaines de ces propriétés. Ces différents Ac sont produits, comme l'a montré
ultérieurement Fagreus en 1948, par des cellules particulières, les plasmocytes qui
appartiennent à l'une des trois grandes catégories de lymphocytes : les lymphocytes B.
L’Immunogène ou l’Antigène se dit donc d'une substance qui, lorsqu'elle est introduite dans
l'organisme, provoque une réaction immunitaire de la part du receveur.
L’Epitope ou déterminant antigénique : c'est la partie de l’antigène qui se lie à l'anticorps.
Les antigènes ont habituellement un grand nombre de déterminants qui peuvent être
différents les uns des autres ou, au contraire, être des structures répétitives.
L'injection à l'animal d'un antigène (Ag) permet d'obtenir un antisérum spécifique de cet Ag,
capable de se combiner spécifiquement avec lui, alors qu'un sérum prélevé chez le même
animal avant son immunisation, est dépourvu de cette propriété ou ne l'exprime que
faiblement. C'est une région particulière de l'Ac, le paratope ou site Ac, qui se lie à une
partie de l'Ag, l'épitope.
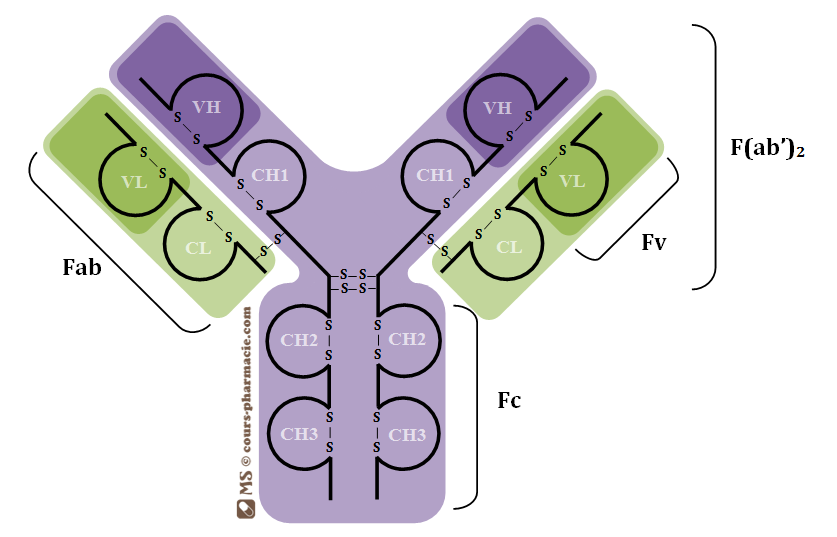
Structure d’un Anticorps

5
Les Ig sont présentes dans le torrent circulatoire et dans certains autres liquides biologiques,
mais aussi à la surface des lymphocytes B qui les synthétisent. Les Ig circulantes et les Ig de
membrane ont une structure légèrement différente, mais toutes les Ig ont une conformation
générale voisine
Ces Ig peuvent également jouer le rôle d'Ag lorsqu'elles sont injectées à un autre animal (de
la même espèce ou d'espèces différentes selon le type de spécificité recherchée). Ainsi, dans
des systèmes hétérologues (par exemple Ig humaines administrées au lapin) les propriétés
antigéniques des Ig ont permis de les répartir en 5 grandes classes d'isotypes : IgG, IgM, IgA,
IgE et IgD avec 4 sous-classes isotypiques pour l'IgG et 2 pour l'IgA.
Une immunoglobuline (Ig) est un anticorps de spécificité inconnue
Un anticorps (Ac) est une immunoglobuline de spécificité connue
Les Anticorps sont des immunoglobulines constituées de glycoprotéines comprenant quatre
chaînes : deux chaînes lourdes identiques et deux chaînes légères identiques réunies entre
elles par des ponts disulfures.
Neuf (9) types de chaînes lourdes : et définissent 9 classes et sous-
classes d'immunoglobulines : IgG, (IgG1, IgG2, IgG3 et IgG4), IgM, IgA (IgA1 et IgA2) IgD et IgE.
Type de Chaîne lourde
Ig correspondant
1
IgG1
2
IgG2
3
IgG3
4
IgG4
IgM
1
IgA1
2
IgA2
IgD
IgE
Les chaînes lourdes peuvent être réunies à 2 types de chaînes légères :et (kappa et
lambda).
Fragment Fab
Fragment Fc
VH = Région variable de la chaîne lourde
CH = Région constante de la chaîne lourde
VL = Région variable de la chaîne légère
CL = Région constante de la chaîne légère
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
1
/
46
100%












{kind=link}