Souris déficientes en chaine invariante - iPubli

NOUVELLES DERNIÈRE HEURE
médecine/sciences
Souris déficientes en chae invariante
a
a
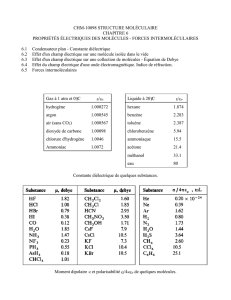
a
s ° 3 vol. 9, rs 93

--
à
T
--
H-2b à
à
à
° 3 vol. rs
--
à
--
à
à
--
--
�
'

8
s ° 3 l. 9, 93
1
/
3
100%