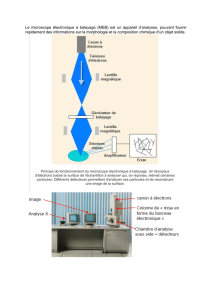
Modélisation de la réponse impulsionnelle pour la détection de

UFR Math´ematiques et Informatique
ED “Cognition, Comportements, Conduites Humaines”
Mod´elisation de la r´eponse impulsionnelle pour
la d´etection de particules en microscopie par
fluorescence
Th`ese de doctorat
soumise le 10 janvier 2008 par
Elodie Dusch
en vue de l’obtention du titre de
Docteur de l’Universit´e Paris Descartes
Composition du jury
Rapporteurs : Pr. Alain Faure Universit´e du Havre
Pr. Isabelle Magnin INSA
Examinateurs : Pr. Nicole Vincent Universit´e Paris Descartes
Pr. Georges Stamon Universit´e Paris Descartes
Pr. Patrick Bouthemy IRISA
Dr. Jean-Christophe Olivo-Marin Institut Pasteur
Dr. Auguste Genovesio Institut Pasteur Korea
Dr. Thierry Dorval Institut Pasteur Korea
Dr. Nicolas Lom´enie Universit´e Paris Descartes


Résumé
La réponse impulsionnelle d’un microscope, appelée Point Spread Function, provoque un flou et une
déformation des objets sur les images de fluorescence. Une connaissance précise de cette fonction peut
permettre de corriger ce phénomène, ou encore de donner un a priori sur la taille des objets recherchés
dans l’image. En effet, sous la supposition que les particules biologiques fluorescentes peuvent être assi-
milées à une impulsion lumineuse, l’image de ces particules sera alors donnée par cette PSF.
Dans un premier temps, nous nous intéressons à la modélisation de la réponse impulsionnelle en
microscopie par fluorescence. Afin d’obtenir un modèle analytique, nous étudions la diffraction des ondes
dans un système optique parfait. Nous proposons un modèle analytique 3D de la PSF d’un microscope
confocal à balayage par ligne en nous basant sur la théorie scalaire de Debye. Ce microscope, dont le
fonctionnement peut être comparé à un microscope à champ large suivant la direction de la ligne et un
microscope confocal classique suivant la direction perpendiculaire, possède une PSF asymétrique dans
le plan focal. La modélisation proposée comprend un modèle asymétrique ainsi qu’une approximation
gaussienne de ce modèle. Une comparaison de ces deux représentations est réalisée sur une vérité terrain.
Dans un deuxième temps, nous utilisons ces modèles pour la détection 3D de particules fluorescentes.
Nous proposons une méthode de détection basée sur la courbure locale, combinée à une modélisation
statistique de la formation de l’image afin d’estimer les positions sous-voxellique des particules détectées.
Cette modélisation prend en compte les perturbations affectant l’image et l’approximation analytique de
la réponse impulsionnelle. De plus, nous étudions l’influence de la modélisation analytique et de son
approximation gaussienne sur la détection de particules sous la résolution optique.
Enfin, nous présentons des applications concrètes de la méthode à des problématiques biologiques,
réalisées en collaboration avec d’autres équipes de l’Institut Pasteur de Corée.

Abstract

Table des matières
Résumé i
Abstract ii
Introduction générale 1
Partie I Modélisation de la réponse impulsionnelle (PSF) 5
Introduction 7
Chapitre 1 Optique ondulatoire 11
1.1 Généralités sur les ondes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.2 Théorie scalaire de la diffraction . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
1.2.1 Principe de Huygens-Fresnels . . . . . . . . . . . . . . . . . . . . . . . . . 13
1.2.2 Théorie de la diffraction de Kirchhoff . . . . . . . . . . . . . . . . . . . . . 14
1.2.3 Approximation de Fraunhofer : modélisation 2D paraxiale . . . . . . . . . . 16
1.2.4 Approximation de Debye : modélisation 3D non-paraxiale/paraxiale . . . . 17
1.2.5 Approche vectorielle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
1.3 Aberrationsoptiques.................................. 19
1.4 Conclusion....................................... 20
Chapitre 2 Réponse impulsionnelle d’un microscope optique à fluorescence 21
2.1 Description des microscopes optiques à fluorescence . . . . . . . . . . . . . . . . . 21
2.1.1 Microscope à champ large . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
2.1.2 Microscope confocal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
2.1.3 CaméraCCD.................................. 23
2.1.4 L’objectif.................................... 24
2.2 ModélisationdelaPSF................................. 25
2.2.1 Microscope optique à champ large . . . . . . . . . . . . . . . . . . . . . . 25
2.2.2 Microscope confocal à balayage par point . . . . . . . . . . . . . . . . . . 26
iii
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
1
/
124
100%