Le système nerveux

1
Le système nerveux
Préparation au concours de kiné 2010
POLYPREPAS

2
les supports anatomiques du message
nerveux
POLYPREPAS
Première partie

3
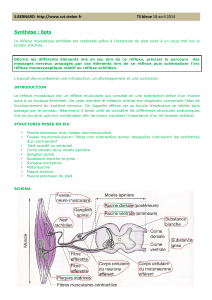
INTRODUCTION
Introduction
Toute réaction de l’organisme à une modification de son milieu extérieur ou de
son milieu intérieur implique une
communication interne. Cela suppose :
• La détection de ce stimulus par des structures spécialisées
• L’émission d’un message transmis directement ou indirectement à des
effecteurs dont les réponses
physiologiques sont généralement adaptées.
Deux types de messages distincts mais complémentaires sont utilisés : le
message nerveux et le message hormonal.

4
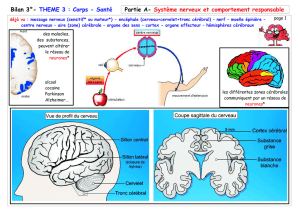
Organisation du système nerveux
I – l’organisation du système nerveux
Le système nerveux de l’homme comme celui des autres vertébrés comprend :
• Des centres nerveux :
Encéphale contenu dans la boite crânienne. La plus grande partie de l’encéphale
est formé par le cerveau constitué des 2 hémisphères cérébraux.
Moelle épinière logée dans le canal rachidien de la colonne vertébrale.
L’ensemble forme l’axe cérébro-spinal.
•Des nerfs : 12 paires de nerfs crâniens reliés à l’encéphale et 31 paires de nerfs
rachidiens rattachés à la moelle épinière.

5
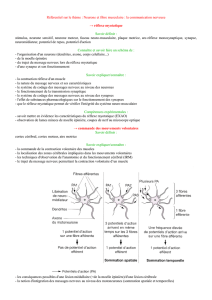
La moelle épinière
A- La moelle
Anatomie
La moelle épinière est un long tube d’environ 1 cm de diamètre logé dans le
canal rachidien constitué par les vertèbres. Elle est formée de la substance
blanche et de la substance grise.
La substance grise contient les corps cellulaires des neurones ainsi que les
prolongements cytoplasmiques des neurones.
La substance blanche contient uniquement des fibres nerveuses c’est à dire des
ramifications des neurones.
La zone dorsale de la moelle est sensitive alors que la zone ventrale est motrice.
Au centre, le canal de l’épendyme prolonge le 4
ème
ventricule de l’encéphale.
La substance grise se répartit en cornes dorsales; cornes intermédiaires et
cornes ventrales.
Les cornes dorsales contiennent des éléments de la sensibilité extéroceptive
(récepteurs situésà la périphérie du corps), de la sensibilité proprioceptive
(récepteurs des muscles, articulations et tendons) et de la sensibilité
viscéroceptive (récepteurs des viscères).
Les cornes ventrales sont composées d’éléments cellulaires à l’origine de la
viscéromotricité (motricité des muscles striés).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
1
/
90
100%