TSSIBG – Les circuits de circulation des lymphocytes et les

TSSIBG – Les circuits de circulation des lymphocytes et les mécanisme moléculaires qui les contrôlent
14/10/2013
MARIANI Natacha L2
TSSIBG
J. Boucrault
10 pages
Les circuits de circulation des lymphocytes et les mécanismes moléculaires qui les contrôlent
A. Introduction
Les lymphocytes sont les cellules qui circulent le plus. Ils circulent :
– Depuis le lieu de leur production (organes lymphoïdes primaires) vers le lieu de leur maturation,
comme le thymus pour les lymphocytes T. Ceci permet l'induction de lymphocytes naïfs.
– Pour rencontrer l’antigène dans les organes lymphoïdes secondaires, où il y a interaction des
lymphocytes T naïfs avec les cellules dendritiques (qui sont aussi recrutées à ce niveau à partir d'un tissu), ce
qui permet l'induction de lymphocytes effecteurs.
– Pour subir les effets d’un environnement cellulaire et cytokinien, c’est la coopération cellulaire
(interactions lymphocytes T – lymphocytes B ; interactions avec les populations non conventionnelles ; … ).
Les lymphocyte est ainsi programmé, et il va ensuite proliférer, être amplifié, puis migrer vers les organes
cibles.
– Pour exercer leurs rôles dans les tissus périphériques : immunosurveillance, fonctions anti-tumorales,
fonctions anti-infectieuses, et surtout fonctions d’homéostasie, de protection et de réparation tissulaire (pour
permettre la réparation, il va y avoir un apport de cellules : c'est l'inflammation).
Les lymphocytes re-circulent. Il y a une mémorisation des informations (antigène, environnement, lieu
dans lequel ça s'est passé...) par les lymphocytes mémoires.
Les autres cellules sanguines circulent également. Les monocytes, qui se différencient en macrophages
dans les tissus, et les cellules dendritiques « immatures ». Ces dernières captent l'antigène puis circulent, mais
ne présentent pas l'antigène (alors que les cellules dendritiques « matures » ne circulent pas mais présentent
l'antigène couplé au CMH).
Les cellules dendritiques maturent au cours de l'inflammation, par exemple.
1/10
Plan
A. Introduction
B. Processus moléculaires guidant les passages à travers les différents tissus
I. Introduction
II. Historique des chimiokines
III. Exemples de processus de migration
IV. Autres paramètres et intérêt du sujet

TSSIBG – Les circuits de circulation des lymphocytes et les mécanisme moléculaires qui les contrôlent
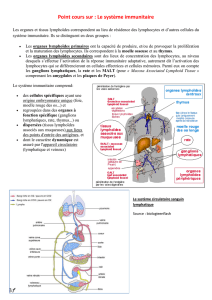
→ Voies de circulation
NB : En ce qui concerne le système nerveux, qui n'a pas de drainage lymphatique, la drainage des antigènes se
fait dans le LCR qui rejoint le système lymphatique.
→ Origine des lymphocytes
2/10

TSSIBG – Les circuits de circulation des lymphocytes et les mécanisme moléculaires qui les contrôlent
Dans la moelle osseuse :
Les cellules souches deviennent des progéniteurs lymphoïdes et des progéniteurs myéloïdes. Les progéniteurs
lymphoïdes se transforment en progéniteurs de lymphocyte T et en lymphocyte B naïfs. Il y a aussi des cellules
dendritiques, plasmocytoïdes ou myéloïdes. Cette classification des cellules dendritiques est morphologique,
car les 2 types peuvent provenir à la fois des progéniteurs lymphoïdes et myéloïdes, et ont les mêmes fonctions.
→ Rencontre entre les lymphocytes et l'information
Le lieu de rencontre est le ganglion lymphatique (organe lymphoïde secondaire).
La cellule dendritique immature va capter l'antigène et va migrer vers le ganglion. Lorsque la cellule
dendritique va être activée (par des cytokines, ….), il va y avoir une activation cellulaire et l'antigène va être
présenté à la surface.
Les progéniteurs lymphocytes T vont dans le thymus où ils vont subir une sélection. En ce qui concerne
l'expression de CD4 et de CD8, les progéniteurs sont soit doubles négatifs (ni CD4, ni CD8), soit doubles
positifs (CD4 et CD8) ou simples positifs (CD4 ou CD8). Après sélection on a des LTCD4 et des LTCD8 dit
naïfs.
Ce LT naïfs vont dans les organes lymphoïdes secondaires (ganglions), où les cellules dendritiques vont leur
présenter l'antigène, pour donner des cellules blastiques qui vont proliférer. Une partie de ces blastes va donner
des LT mémoires qui ont une longue durée de vie.
Les cellules blastiques vont aller dans les organes périphériques et pour se transformer en LT effecteurs. De
même que précédemment, une partie de ces LT effecteurs va devenir des LT effecteurs mémoires qui
survivront longtemps. Le devenir majoritaire des LT effecteurs est de mourir (apoptose).
Les LT mémoires, qui eux survivent, vont circuler et regagner les organes lymphoïdes secondaires et la moelle
osseuse, ce qui permettra une réponse plus rapide lors d'une présentation ultérieure du même antigène.
LB sort de la moelle osseuse à l'état de LB naïf. Leur devenir est de devenir des plasmocytes qui produiront les
anticorps. Les LB ont cependant des anticorps de surface qui leur permettent de reconnaître l'antigène
directement (contrairement aux LT qui ont besoin d'un contact avec une autre cellule pour reconnaître
l'antigène).
Le LB « naïf » se rend dans les organes lymphoïdes secondaires où l'antigène lui est présenté pour qu'il se
transforme en blaste. Dans la plupart des cas, cette transformation nécessite un « help » des cellules T. Cette
coopération cellulaire entre LB et LT nécessite la présence des cellules dendritiques.
On a ensuite une réponse immunitaire, avec la production de plasmocytes qui vont circuler puis se retrouver
dans les organes lymphoïdes secondaires ou dans les tissus périphériques.Les LB mémoires re-circulent et
peuvent se retrouver par exemple dans la moelle osseuse.
3/10

TSSIBG – Les circuits de circulation des lymphocytes et les mécanisme moléculaires qui les contrôlent
→ Il y a un autre niveau de circulation : circulation au sein des organes
La répartition des cellules est organisée dans les organes lymphoïdes
=> l'inflammation modifie la composition et la répartition des différents types cellulaires.
→ communication entre les cellules
Les cellules circulent mais rien ne se passe si elles ne communiquent pas. Il y a donc formation de contacts
étroits entre 2 types cellulaire, donc formation de synapses.
Les interactions cellulaires sont organisées en synapses.
Il existe 2 types de synapses : les synapses nerveuses et les synapses immunitaires (très similaires aux synapses
nerveuses).
La différence la plus importante entre les 2 systèmes (nerveux et immunitaire) est la circulation.
4/10

TSSIBG – Les circuits de circulation des lymphocytes et les mécanisme moléculaires qui les contrôlent
On peut caractériser la synapse immune de différentes façons. Par sa qualité, par exemple (temps de contact,
force de contact, adhésion), qui va faire qu'on aura ou non une réponse immunitaire.
Ex : durée de contact entre les cellules dendritiques et les LT.
Si le contact est rapide (cellules dendritiques I-CAM négatives), il y aura présence de LT
cytotoxiques mais pas de LT mémoires, alors que si le contact est prolongé (cellules dendritiques I-CAM
positives) on aura des LT cytotoxiques et des LT mémoires.
Donc, l'étreinte prolongée est nécessaire pour installer une mémoire efficace de la réponse
immunitaire.
B. Processus moléculaires guidant le passage dans les différents tissus
I. Introduction
La circulation lymphocytaire est un événement majeur de la réponse immunitaire pour leur maturation, le
développement de la réponse immunitaire et pour qu'un lymphocyte, induit par une « information » tissulaire,
soit dirigé préférentiellement vers le tissu concerné.
C'est un processus programmé et lié à l'expression constitutive ou induite :
- de panels de molécules d'adhérence exprimées par les lymphocytes et les cellules qu'ils contactent
ex : I-CAM1
- de panel de récepteurs à 7 domaines TM
→ récepteur aux chimiokines
→ autres récepteurs : récepteurs au C3a, C5a, récepteur de la sphingosine-1-phosphate
- de protéases matricielles qui favorisent leur migration (les protéases lysent l'environnement pour permettre la
migration des cellules).
II. Historique des chimiokines
Le lipopolysaccharide (LPS) induit la libération par les monocytes de l'interleukine-8 qui attire les
neutrophiles. (IL8 est en fait une chimiokine, ce qui a été découvert plus tard, donc cette molécule a plusieurs
noms, comme beaucoup d'autres molécules en immunologie). Depuis, la famille comprend plus de 50 ligands
qui interagissent avec au moins 18 récepteurs. Ce sont des récepteurs à 7 domaines transmembranaires associés
aux petites protéines G. Les chimiokines sont des petites molécules, très basiques (ce qui leur permet d'interagir
avec les protéines très acides de la matrice extracellulaire, et on a donc un gradient de chimiokines dans la
MEC), de 8 à 10 kD. Leur interaction avec des protéines comprenant des groupements sulfates et les
protéoglycans entraîne la formation d'un gradient.
→ Migration des cellules grâce aux chimiokines :
Au niveau du centre de migration de la cellule, on a une concentration de récepteurs aux chimiokines, ce qui
induit une activation de la matrice extracellulaire. On aura ensuite la polarisation du cytosquelette et ensuite une
contraction. À l'uropode, on n'a pas de récepteurs, mais il y a des molécules anti-adhésives qui vont faciliter la
dé-adhésion et l'avancée.
Ce qui fait la migration des cellules est donc la mise en place du gradient de chimiokines et la polarisation
cellulaire.
5/10
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%