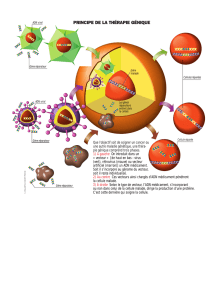
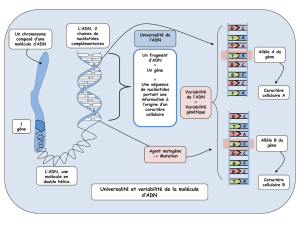
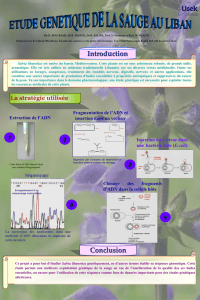
Septembre 2009

Septembre 2009
Techniques de biologie moléculaire
Mme Bonnet
I. La séparation des composants des génomes
La plupart des EV ont des génomes fragmentés en composants distincts.
Bact : En plus de leur ADN Xmique, elles possèdent un plasmide (molécule circulaire double
brin) Elles possèdent également des génomes au niveau des chlp et des mito.
A. Séparation par leur taille
Les fragments d’ADN de faible taille (<50kb) peuvent être séparer par électrophorèse sur un
gel, qui permet de faire un tamisat moléculaire
Contrairement aux prot dont la charge est variable, tous les AN ont une charge négative
équivalente => leur migration sur un gel va être uniquement fonction de leur taille.
2 types de matériau pour un gel :
- acrylamide
- agarose
Si on utilise une faible concentration de ces matériaux, on pourra séparer des fragments de
grande taille.
A l’inverse, si on utilise une forte concentration, on ne pourra séparer les fragments que de
petite taille (plus la concentration est forte, plus les mailles sont serrer, plus les petites
passeront comparer aux grosses)
Si on utilise une faible [C] en acrylamide, le gel ne sera pas assez solide, on n’utilise ce
matériau que pour analyser les petites molécules. En général, on ne l’utilise que pour le
séquençage.
A l’inverse l’agarose, ayant une rigidité mécanique bien supérieure, on peut l’utiliser à très
faible concentration, on peut l’utiliser à très faible concentration pour des molécules de 100
pb à 50 kb.
On utilise le sybersafe au lieu du bromure d’éthidium (BET)
BET : molécule très dangereuse : c’est un mutagène très fort.
Cette molécule, appelée aussi intercalant se
place entre les bases de l’ADN et après
électrophorèse et UV, on retrouve
l’intercalant sous forme fluorescente, ce
qui permet de voir où les AN se sont
déposés.
Pour l’ARN c’est quasiment la même chose. Mais il va falloir utiliser des conditions
dénaturantes pour éviter que la molécule d’ARN ne se replie sur elle-même.
Il faut que la forme des ARN soit identique
si on veut pouvoir les séparer.

On utiliser les molécules suivantes :
- méthylmercure
- formaldéhyde
Rappel : électrophorèse
Support horizontal, on fait passer un
courant électrique de (-) vers (+).
Les molécules migrent en fonction de leur
taille. Les petites vont migrer plus
facilement.
Les bandes ne sont visibles que sous UV.
B. Séparation par leur forme
On va pouvoir séparer des molécules de
même taille en fonction de leur degré de
surenroulement
Un plasmide en solution adopte 3 formes
différentes, on peut retrouver ces 3 formes
sur un gel d’agarose :
Séparation de molécules de densités différentes
On utilise un gradient de densité de BET.
On va utiliser du chlorure de césium
(CsCl) pour obtenir des solutions de très
forte densité allant jusqu’à 1,9.
Dans un tube, celé, on met CsCl + AN +
BET.
On fait une ultracentrifugation à 500 000 g
pendant plusieurs heures.
Un force centrifuge va avoir tendance à
entraîner les molécules de forte densité au
fond du tube.
Il va se créée un gradient de densité, avec au fond une densité de 1,9 et en haut une densité de
1. Ce gradient est stable une fois que la centrifugation est finie.
En passant le tube sous UV on va observer différentes bandes :
- ARN à d = 1,9
- ADN à d = 1,7
- Prot à d = 1,6
Pour ne prélever que les molécules
d’ADN, on utilise une seringue.

C. Séparation par leur composition en bases
Incidence sur la densité des molécules.
Les ADN étant riches en G et C seront plus denses que ceux riches en A / T. Ceci lié au
nombre de liaison entre les bases.
II. Les techniques de transfert
Une fois qu’on a réussi à séparer des
molécules sur un gel d’électrophorèse, on
se retrouve avec tout un ensemble (des fois
avec un génome entier) mais on ne se sait
pas à quelle bande correspond quel gène.
Il faut pouvoir détecter la présence d’un
seul gène parmi toutes les bandes présentes
sur le gel.
Pour y arriver, on va réaliser un transfert
pour nous permettre d’obtenir sur un filtre
de nitrocellulose ou de nylon (feuille très
fine et très fragile) un seul gène visible.
A. Le transfert de l’ADN
= Southern blotting ou Southern blot
Préparation au transfert :
- On fait migrer de l’ADN sur un gel d’agarose
- On fait traiter le gel d’électrophorèse par de l’acide chlorhydrique => dépurination
limitée de l’ADN (élimination des purines (A/G) de façon partielle). La molécule se
retrouve morcelée.
- Le gel va être placer dans une solution alcaline en présence de soude qui va entraîner
une dénaturation partielle, elle va créer des coupures simple brin au niveau des trous
créer auparavant.

L’ADN fixé sur le gel peut aller se fixer plus facilement sur le filtre.
Transfert :
- On laisse 12h et à Tre ambiante.
- On obtient un flux ascendant vertical,
toutes les molécules passent du gel au
filtre.
Toutes les molécules d’ADN ont été
transférées mais on ne les voit pas.
- Pour mettre en évidence la présence
d’un gène donné, on va utiliser des
sondes moléculaires (molécules d’AN
dont on connaît la séquence marquées
avec un atome radioactif le 32P,
complémentaires d’une partie du gène
qu’on veut détecter, il faut que le gène
soit simple brin).
Sonde radioactive : sonde chaude.
Sonde non radioactive : sonde froide.
- Dans une cuve avec une sonde, on met
le filtre et on agite à environ 65 °C
pendant plusieurs heures. Ce qui
entraîne des collisions entre l’ADN et
les sondes. Quand le gène et la sonde
se rencontre, il va y avoir interaction.
- On espère qu’une sonde s’est fixée.
Lavage, agitation.
- On met le filtre au contact d’une radio
dans une boite. ON met le tout à –40°C
pendant plusieurs heures. L’atome * va
émettre des rayons β et marquer la
molécule.
- Sur la radio, on va observer une tache noire à l’emplacement du gène. Si il n’y a pas de
tache, soit le gène n’est pas présent, soit la manipulation n’a pas marchée.
Il existe 2 types de sonde :
- Sonde hétérologue : provenant d’une autre espèce d’où on a extrait l’ADN
La sonde n’est pas forcément complémentaire à 100% du gène, hybridation pas à
100%, présence de boucle entre la sonde et le gène.
- Sonde homologue : provenant de la même espèce
Hybridation totale, plus difficile à réaliser, du fait du nombre de liaison à établir. Pour
faciliter cette hybridation, on va augmenter la température ce qui augmente le nombre
de collision entre les 2 molécules et augmente les chances de rencontre et l’hybridation
pourra se faire.
Cette hybridation est beaucoup plus spécifique.

B. Le transfert de l’ARN
= Northern Blot
Mettre en évidence de l’expression d’un gène.
L’ARN on le fait migrer sur un gène d’agarose en conditions dénaturantes puis on passe
directement au transfert.
On laisse agir toute la nuit à Tre ambiante.
Il y a eu un flux descendant qui a entrainé les molécules d’ARN sur le filtre de nitrocellulose.
On met en contact avec la sonde, puis des lavages, puis une autoradiographie.
Sur l’autoradiographie, on devrait obtenir une bande (expression du gène). S’il n’y en a pas, le
gène ne s’est pas exprimé dans les conditions dans lesquelles on a extrait l’ARN, le gène n’est
pas présent ou la manipulation n’a pas marché.
C. Le transfert des protéines
= Western Blot
On fait migrer les protéines sur un gel d’électrophorèse d’acrylamide avec du SDS (sodium
dodecyl sulfate). Elles ont la même charge et donc vont migrer uniquement en fonction de
leur taille.
Le transfert :
A la vertical, gel + filtre en milieu liquide dans une cassette, on laisse à 4 °C pendant quelques
heures, les molécules se déplacent sur le filtre.
On met en présence spécifique à la protéine.
Lavages
Autoradiographie => bande => présence de la protéine, expression du gène.
Pas de bande => pas d’expression du gène.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
1
/
26
100%